Origine degli eucarioti. Teoria simbiotica, endosimbiosi o simbiogenesi Teoria simbiotica
Va notato tre ipotesi per l'origine delle cellule eucariotiche:
- ipotesi simbiotica, O simbiogenesi,
- intussuscezione,
- chimerico.
Oggi nel mondo scientifico viene riconosciuta l'ipotesi principale sull'origine degli eucarioti simbiogenesi.
Secondo la simbiogenesi, tali organelli di cellule eucariotiche come mitocondri, cloroplasti e flagelli sono nati attraverso l'introduzione di alcuni procarioti in un'altra cellula procariotica più grande, che ha svolto il ruolo di cellula ospite.
L'ipotesi simbiotica ha difficoltà nello spiegare l'origine del nucleo delle cellule eucariotiche e nella questione di quale procariota abbia agito come ospite. I dati dell'analisi molecolare del genoma e delle proteine degli eucarioti mostrano che, da un lato, si trattava di un organismo vicino agli archaea (precedentemente classificati come batteri, quindi erano separati in un ramo separato). D'altra parte, gli eucarioti contengono proteine (e geni responsabili della loro sintesi) caratteristici di gruppi completamente diversi di procarioti.
Secondo l'ipotesi dell'invaginazione dell'origine delle cellule eucariotiche, i loro organelli si sono formati mediante invaginazione della membrana citoplasmatica con successiva separazione di queste strutture. Si formarono qualcosa come palline, circondate da una membrana e contenenti all'interno il citoplasma e composti e strutture qui catturati. A seconda di ciò che veniva ingerito si formavano diversi organelli.
I procarioti non hanno veri e propri organelli; le loro funzioni sono svolte da invaginazioni di membrana. Pertanto, è facile immaginarne un simile slacciamento. A favore dell'ipotesi dell'invaginazione c'è anche la somiglianza della membrana citoplasmatica e delle doppie membrane degli organelli.
Dal punto di vista dell'invaginogenesi, l'origine del nucleo è facilmente spiegabile, ma è inspiegabile il motivo per cui il genoma e i ribosomi del complesso nucleare-citoplasmatico differiscono da quelli dei cloroplasti e dei mitocondri (ricordiamo che hanno anche DNA e ribosomi). Inoltre, in questi organelli, il sistema di biosintesi delle proteine (DNA, RNA, ribosomi) è simile a quello dei procarioti.
Questa differenza è ben spiegata proprio dal punto di vista della prima ipotesi, quella simbiotica. Secondo esso, il procariota anaerobico in qualche modo è entrato nel procariota aerobico. Non è stato digerito, ma, al contrario, ha iniziato a nutrirsi a spese della cellula ospite. A sua volta, ha utilizzato l'ossigeno per ottenere energia, e questo metodo di ossidazione è molto più efficiente; ha ceduto l'energia in eccesso all'ospite procariotico, che anche in questo caso ne ha beneficiato. Si è creata una simbiosi. Successivamente, il procariota invasore si semplificò, parte del suo genoma migrò nella cellula ospite e non poté più esistere in modo indipendente.
La simbiogenesi spiega in modo simile l'origine dei cloroplasti. Furono introdotti solo i procarioti capaci di fotosintesi (simili alle alghe blu-verdi).
I primi eucarioti senza simbionti fotosintetici diedero origine agli animali in cui apparvero: le piante.
Attualmente esistono protozoi (organismi eucariotici unicellulari) che non hanno mitocondri né cloroplasti. Ma invece di loro, i procarioti simbionti si stabiliscono nel citoplasma e svolgono le funzioni corrispondenti. Questo fatto, così come la somiglianza del sistema di biosintesi proteica dei mitocondri e dei plastidi con i procarioti, sono considerati una prova della simbiogenesi. La prova viene anche dal fatto che mitocondri e cloroplasti si riproducono in modo indipendente; non vengono mai costruiti ex novo dalla cellula.
La terza ipotesi, chimerica, sull'origine delle cellule eucariotiche è supportata dalle grandi dimensioni del loro genoma, che supera migliaia o più volte il genoma batterico, nonché dalla diversità delle proteine sintetizzate presenti in diversi gruppi di procarioti. È chiaro che durante l'evoluzione degli eucarioti, il loro genoma è diventato più complesso, è raddoppiato e in esso sono comparsi molti geni regolatori. Tuttavia, l’aumento iniziale delle dimensioni del genoma potrebbe essere dovuto alla combinazione dei genomi di diversi procarioti.
Forse nell'antichità un certo procariota acquisiva la capacità di fagocitare e, nutrendosi in questo modo, assorbiva altri procarioti che non sempre venivano digeriti. Il loro genoma conteneva geni utili all'ospite e lui li incluse nel suo genoma. Forse alcuni dei procarioti finiti all'interno sono diventati organelli, il che combina l'ipotesi chimerica con la simbiogenesi.
Poiché tutte le ipotesi di cui sopra hanno punti di forza e di debolezza e per molti versi non si escludono a vicenda, quindi, a nostro avviso, una combinazione di molti fattori descritti da diverse ipotesi potrebbe aver avuto un ruolo nell'origine delle strutture delle cellule eucariotiche .
Va inoltre notato che secondo l'ipotesi simbiotica, l'origine di altri organelli di membrana come i vacuoli, il complesso di Golgi, ecc. può essere considerata come un'ulteriore semplificazione, ad esempio i mitocondri.
Notiamo anche che la combinazione di proteine di diversi procarioti negli eucarioti potrebbe non indicare affatto il chimerismo dei primi. È possibile che alcune funzioni enzimatiche possano essere eseguite solo da proteine con una struttura specifica. E l'evoluzione degli eucarioti è arrivata ripetutamente a questo punto, indipendentemente dall'evoluzione di gruppi di procarioti che non erano i loro antenati. Diciamo che c'era convergenza a livello molecolare.
I mitocondri sono fedeli compagni degli eucarioti. Secondo la teoria della simbiogenesi, è stata l'acquisizione dei mitocondri a provocare la formazione degli organismi nucleari. Una delle prove di questa teoria fu la scoperta di mitocondri o organelli simili in tutti gli eucarioti, anche i più semplici. Ma nel maggio 2016, un team di scienziati cechi ha descritto il primo organismo nucleare della storia che non conteneva nemmeno segni indiretti di mitocondri. Questa scoperta potrebbe scuotere le idee attuali sull’evoluzione iniziale degli eucarioti?
I mitocondri sono organelli a doppia membrana che forniscono energia alle cellule di quasi tutti gli eucarioti. È noto con certezza che sono imparentati con gli α-proteobatteri e sono entrati a far parte delle cellule eucariotiche circa 1,5 miliardi di anni fa. L'origine batterica dei mitocondri è evidenziata dalla presenza di due membrane (interna ed esterna dell'ospite), dal proprio DNA circolare e dalla macchina di traduzione, nonché dalla capacità di dividersi in modo indipendente. Alcuni credono addirittura che l'apoptosi sia un tentativo dei mitocondri di uccidere l'utilizzatore che l'ha assorbito.
Tuttavia, non c'è ancora consenso nella comunità scientifica riguardo al ruolo di questa simbiosi nello sviluppo degli eucarioti (Fig. 1). Sostenitori Teorie della simbiogenesi sostengono che la fusione di un certo archeobatteri con l'antenato dei mitocondri (batteri) ha lanciato una catena di eventi che hanno portato alla formazione dei moderni eucarioti. Seguaci teoria archeozoica (ipotesi) Al contrario, credono che solo un organismo nucleare già formato, un archezoo, possa “domare” il mitocondrio.
Grazie a Carl Woese e George Fox, che negli anni '70 del secolo scorso confrontarono i geni 16S rRNA di molti esseri viventi, cosa che per lungo tempo aveva ingannato i microbiologi con la loro somiglianza visiva, le due forme di organismi prenucleari (procarioti) furono separate completamente e irrevocabilmente , e anche al livello più alto: gli eubatteri sono stati privati del loro diritto alla "verità" (-eu), gli archeobatteri hanno il diritto di portare l'orgoglioso nome di batteri, ma nel nuovo sistema di organismi viventi è stato loro assegnato il proprio dominio ( taxon di rango più elevato): “ L'evoluzione tra l'incudine e il martello, o come la microbiologia ha salvato l'evoluzione dall'assorbimento della biologia molecolare" E " Carl Woese (1928–2012)". Così nel 1990, è stato chiesto a una persona di rendersi conto che tutti gli esseri viventi sono filogeneticamente divisi in tre domini: batteri, archaea ed eucarioti, e i batteri differiscono dagli archaea ancor più che dagli archaea dagli eucarioti, e recentemente hanno quasi dubitato dell'opportunità di dividere questi ultimi : " Trovati gli antenati di tutti gli eucarioti". Tuttavia, la proposta di quasi trent'anni fa non ha ancora trovato comprensione tra molti autori di libri di testo di biologia domestica. E cosa? E se questi scienziati cambiassero di nuovo tutto e dovessero riscrivere i libri di testo ogni vent’anni o qualcosa del genere? - Ed.
Nel 1928, il famoso biologo della fine del XIX e XX secolo, Edmund Wilson, parlò dell'ipotesi dell'origine batterica dei mitocondri come segue: “Tali idee sono troppo fantastiche per essere discusse in una società biologica decente”.. Oggi, un atteggiamento simile si è formato nei confronti della teoria archeozoica e il ruolo chiave dei mitocondri nell'evoluzione iniziale degli eucarioti è generalmente riconosciuto. Apertura del primo vero protozoo non mitocondriale ti fa riflettere nuovamente sui punti di forza e di debolezza di ciascuna teoria.
Teoria della simbiogenesi
Uno dei vantaggi della teoria simbiogenetica rispetto alla teoria degli archeozoi è che spiega l'emergenza del nucleo e la struttura intronica del genoma. Nei procarioti è molto diffuso il trasferimento genico orizzontale (HGT), grazie al quale le popolazioni possono scambiarsi rapidamente parti del pool genetico. L'HGT contribuisce all'insicurezza del genoma: dopo tutto, il DNA in arrivo in questo caso non è in alcun modo separato dal contenuto della cellula ospite.
Molto probabilmente i tentativi da parte della cellula ospite di distruggere il simbionte non ancora addomesticato hanno portato al rilascio di DNA simbiotico nel citoplasma. Questo DNA, trovandosi in prossimità del genoma ospite, potrebbe facilmente integrarsi in esso. A causa dell'HGT, anche negli eucarioti che hanno perso i mitocondri, inizialmente vengono trovati i geni mitocondriali.
Una tale fusione di genomi potrebbe, in primo luogo, contribuire allo sviluppo dell’interdipendenza tra il simbionte e l’ospite. In secondo luogo, un abbondante HGT potrebbe trasportare non solo geni che assicuravano l'intreccio del metabolismo di due organismi, ma anche retroelementi egoistici. L'invasione degli introni del gruppo II, sfuggiti agli α-proteobatteri, ha portato all'allentamento del genoma dell'ospite inizialmente molto denso: fino all'80% del DNA dell'ospite era ora costituito da introni. In una situazione così complessa, la cellula ospite ha sviluppato diverse linee di difesa per il suo genoma da una raffica di introni: un sistema di membrane interne e un nucleo, un sistema di ubiquitina per la degradazione delle proteine danneggiate, decadimento dell'RNA mediato da sciocchezze e altri sono emerse le caratteristiche caratteristiche degli eucarioti (Fig. 3).
Figura 3. Formazione delle caratteristiche principali degli eucarioti può essere spiegato dall'invasione degli introni del gruppo II nel genoma della cellula ospite che ha seguito la simbiosi mitocondriale.
Un'altra potente prova della simbiogenesi è il fabbisogno energetico degli eucarioti. Sebbene il consumo energetico dei pro- e degli eucarioti per grammo di peso sia approssimativamente lo stesso, le cellule nucleate sono molto più grandi delle cellule non nucleate, inducendole a consumare circa 5000 volte più energia (2300 pW/cella contro 0,5 pW/cella). Quando si ricalcola il consumo di energia per il gene medio di un organismo unicellulare, si scopre che un gene eucariotico consuma 1000 volte più energia. Senza l'energia mitocondriale sarebbe impossibile non solo creare organismi complessi, grandi e in movimento attivo, ma anche garantire il funzionamento delle strutture cellulari tipiche degli eucarioti.
Nei batteri giganti, il ridimensionamento dell'energia procariotica dovuto alla poliploidizzazione di massa (come nel caso Epulopiscium, crescendo fino a 0,6 mm e contenente 200.000 copie di un genoma che misura 3,8 milioni di bp) non porta ad un aumento della produzione di energia per gene e la cellula rimane tipicamente batterica. - Auto.
Un altro fatto importante a sostegno dello scenario simbiogenetico è l'esistenza di simbionti batterici intracellulari. I casi di endosimbiosi nei batteri sono estremamente rari in natura, ma esistono ancora e dimostrano come potrebbe formarsi il dominio della vita eucariotica.
Teoria archeozoica
Gli archeozoi sono i presunti antenati non mitocondriali ma nucleari dei moderni eucarioti. Secondo lo scenario archeozoico, i mitocondri furono addomesticati solo nelle ultime fasi dell'evoluzione eucariotica e non ebbero un impatto significativo su questo processo.
Una delle principali disposizioni della simbiogenesi è l'ipotesi della semplicità iniziale. Si sa molto poco della vita durante il Proterozoico, quindi ci sono molte ipotesi spesso mutuamente esclusive sulla sua struttura. Se, secondo la prima ipotesi, si ritiene che eucarioti più complessi si siano evoluti da procarioti con genomi molto compatti, allora nello scenario archeozoico inizialmente esistevano cellule con genomi confusi e voluminosi, da cui si sono evoluti per riduzione i procarioti più semplici. Gli eucarioti conservavano solo la loro complessità primaria.
L'evoluzione dei genomi non sempre passa da semplice a complessa. E tra gli eucarioti ci sono esempi che lo confermano.
Tuttavia, la riduzione del genoma non accompagna necessariamente la sua compattazione. La prova di ciò può essere trovata sia nelle forme di vita protozoiche che multicellulari.
Ad esempio, i ciliati a vita libera Paramecio tetraurelia contiene 30.000 geni, ciascuno dei quali ha una media di 2 kb. Questa compattezza si ottiene riducendo la dimensione degli introni al massimo di 25 bp. e riducendo le distanze intergeniche.
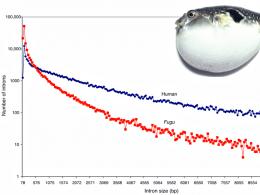
Anche i vertebrati possono avere genomi insolitamente compatti: il genoma del pesce palla è otto volte più piccolo di quello umano, in gran parte a causa del suo basso contenuto di ripetizioni (Figura 4).

Figura 4. Il pesce palla ha un genoma insolitamente compatto per un vertebrato. in parte a causa degli introni corti. L'asse verticale è riportato su scala logaritmica.

Figura 5. Ipotesi di complessità iniziale implica che i rami della vita procariotica si siano evoluti da forme più complesse attraverso la riduzione. Il vettore di riduzione dello sviluppo potrebbe essere stato impostato dai primi archeozoi predatori, opprimendo altri organismi.
Gli esempi sopra riportati mostrano che la semplicità dei genomi procariotici può risultare secondaria. Se questo è vero, allora LUCA – l’ultimo antenato comune di tutti gli organismi moderni – potrebbe avere un genoma eucariotico.
L'ipotesi della complessità iniziale è confermata anche dai cosiddetti geni caratteristici (“firme”) - geni eucariotici che non hanno omologhi procariotici. Molto probabilmente, questi geni erano contenuti in LUCA, ma furono persi da batteri e archaea.
Sfortunatamente per la teoria archezoica, dall’inizio del 21° secolo l’elenco delle firme si è notevolmente assottigliato. Tra i tanti genomi sequenziati da allora, sono stati trovati i loro omologhi procariotici. Pertanto, ogni anno compaiono sempre più proteine, la cui presenza negli eucarioti può essere spiegata dal fatto che i loro geni sono stati portati da un antenato archeologico o batterico durante la simbiogenesi.
E allo stesso tempo, la scoperta di omologhi procariotici delle proteine del movimento cellulare (actine, tubuline e chinesine) conferma indirettamente la possibilità che gli archeozoi possano muoversi attivamente e persino essere i primi predatori sulla Terra capaci di fagocitosi. L'emergere di predatori nella comunità autotrofa-saprotrofica della culla della vita avrebbe dovuto avere un impatto colossale sul corso dell'evoluzione. Negli scenari più entusiasmanti, alcune prede archezoiche si adattano alla rapida divisione e crescita, mentre altre si adattano a nicchie in cui l'archezoo non può penetrare. Di conseguenza, l'ipotetico archezoo guidò l'evoluzione dei suoi contemporanei lungo un percorso riduttivo con un'enfasi sulla flessibilità metabolica e sulla velocità di divisione, durante la quale si formarono i procarioti a noi noti (Fig. 5).
Ma nonostante il fatto che la teoria archeozoica abbia alcuni punti forti o almeno sferri colpi mirati alla simbiogenesi, manca la cosa principale: non spiega come o perché si sia formato il nucleo.
Trovata unica
Negli anni ’80 c’erano molti contendenti al titolo di archeozoi moderni, ma negli anni successivi in tutti furono trovati organelli simili ai mitocondri (mitosomi e perossisomi) e geni marcatori del passato mitocondriale: geni per l’assemblaggio di Fe- Proteine S, trasportatori e chaperoni mitocondriali, cardiolipina sintetasi. Inoltre, alcune proteine sintetizzate nel citoplasma hanno sequenze importate nei mitocondri, che possono persistere in assenza dei mitocondri stessi.
Con ogni nuova "chiusura" di un potenziale archeozoo, lo scenario non mitocondriale per la formazione degli eucarioti si è rivelato sempre meno probabile. E poi, nel maggio 2016, è finalmente apparso un nuovo potenziale archezoico, che non conteneva nemmeno una traccia di mitocondri.
Stiamo parlando di ossimonato anaerobico Monocercomonoidi sp. PA203, che vive nell'intestino degli insetti. Gli ossimonadi sono privi di mitocondri e non contengono geni di origine mitocondriale nel loro DNA nucleare. Ricevono energia dalla glicolisi che avviene nel citoplasma.
Genoma Monocercomonoidi sp., decifrato da un team di scienziati cechi, contiene 16.629 geni, tra i quali non ci sono i marcatori sopra menzionati. Anche la ricerca di omologhi mitocondriali e di proteine con sequenze di importazione non ha dato risultati soddisfacenti (Fig. 6).
L'unica cosa che è stata trovata sono stati due geni, i cui prodotti sono stati trovati in un parente stretto Monocercomonoidi sp. possono (o meno) essere contenuti nei mitocondri, ma mancano di sequenze di importazione.
Lo credono gli autori della scoperta Monocercomonoidi una volta contenevano mitocondri, poiché generi strettamente imparentati hanno tracce di mitocondri. È ancora possibile che questi protozoi abbiano mitosomi non ancora rilevati che sono diventati così degradati che non è rimasta alcuna prova della loro presenza nel genoma.
Comunque, Monocercomonoides sp.- finora un caso unico di protista veramente privo di mitocondri nell'intera storia della biologia. E questo caso dimostra che gli eucarioti possono vivere non solo senza mitocondri, ma anche senza il loro patrimonio genetico.
In quale sequenza gli antenati degli eucarioti acquisirono proprietà intracellulari e quale divenne il biglietto fortunato per il futuro evolutivo, potete scoprirlo dall'articolo “ La genealogia delle proteine suggerisce l'acquisizione tardiva dei mitocondri da parte degli antenati eucariotici» . - Ed.
Questa scoperta, ovviamente, non infligge un duro colpo alla teoria della simbiogenesi, ma solleva sicuramente interrogativi su cosa è necessario e cosa è in eccesso negli eucarioti.
Letteratura
- Come sono comparsi i mitocondri (una storia simile a una fiaba);
- Kunin E.V. Logica del caso. M.: Tsentrpoligraf, 2014. - 527 pp.;
- L'evoluzione tra l'incudine e il martello, o come la microbiologia ha salvato l'evoluzione dall'essere inghiottita dalla biologia molecolare;
- Carl Woese (1928–2012);
- Kondratenko Y. (2015). "Abbiamo trovato gli antenati di tutti gli eucarioti." "Il gatto di Shroedinger". 6 ;
- van der Giezen M. (2009).
Cinquant'anni fa, nel 1967, Lynn Margulis pubblicò un ampio resoconto della teoria simbiogenetica, secondo la quale gli eucarioti (organismi con nuclei cellulari) sorsero come risultato di una serie di associazioni di cellule diverse tra loro. Un emendamento moderno a questa teoria afferma che la formazione degli eucarioti, apparentemente, non fu una tendenza generale che abbracciava molti rami evolutivi (come supponeva Margulis), ma un evento unico che portò alla fusione di cellule arcaiche e proteobatteriche. Di conseguenza, si formò una cellula complessa con mitocondri, che divenne il primo eucariota. Ulteriori eventi simbiogenetici - ad esempio la cattura di alghe che divennero cloroplasti - si sono verificati molte volte, ma non sono associati alla comparsa degli eucarioti in quanto tali.
Più di cinquant'anni fa, nel marzo 1967, il Journal of Theoretical Biology pubblicò un articolo “Sull'origine delle cellule che si dividono per mitosi” (L. Sagan, 1967. Sull'origine delle cellule mitosanti). L'autore dell'articolo si chiamava Lynn Sagan, ma questa donna straordinaria in seguito divenne molto più conosciuta come Lynn Margulis. Portava il cognome Sagan perché fu brevemente sposata con Carl Edward Sagan, astronomo e scrittore.
La pubblicazione di un articolo di Lynn Margulis nel 1967 (lo chiameremo così per comodità) segnò l'inizio di un rinnovamento dei concetti biologici, che molti autori considerarono un cambio di paradigma, cioè una vera e propria rivoluzione scientifica. (I.M. Mirabdullaev, 1991. Teoria endosimbiotica - dalla finzione al paradigma). L'essenza dell'intrigo qui è semplice. Sin dai tempi di Charles Darwin, i biologi sono convinti che il principale metodo di evoluzione sia la divergenza: la divergenza dei rami. Lynn Margulis fu la prima a spiegare veramente alla comunità scientifica che il meccanismo di alcuni importanti eventi evolutivi era probabilmente fondamentalmente diverso. Gli interessi di Margulis si sono concentrati sul problema dell'origine degli eucarioti, organismi le cui cellule hanno una struttura interna complessa con un nucleo. Gli eucarioti comprendono animali, piante, funghi e molti organismi unicellulari: amebe, flagellati, ciliati e altri. Margulis ha dimostrato che l'evoluzione iniziale degli eucarioti non si riduceva affatto alla divergenza: includeva la fusione di rami evolutivi, e più di una volta. Il fatto è che almeno due tipi di organelli eucariotici - i mitocondri, grazie ai quali possiamo respirare ossigeno, e i cloroplasti, che svolgono la fotosintesi - non provengono dallo stesso antenato della parte principale della cellula eucariotica (Fig. 1) . Sia i mitocondri che i cloroplasti sono ex batteri, inizialmente completamente estranei agli eucarioti (proteobatteri nel caso dei mitocondri e cianobatteri nel caso dei cloroplasti). Questi batteri furono assorbiti dalla cellula di un antico eucariota (o antenato degli eucarioti) e continuarono a vivere al suo interno, conservando per il momento il proprio apparato genetico.
Pertanto, una cellula eucariotica è, come dice Margulis, sistema multigenomico. Ed è nato come risultato della simbiosi, cioè della convivenza reciprocamente vantaggiosa di diversi organismi (più precisamente, dell'endosimbiosi, uno dei partecipanti che vive all'interno dell'altro). I rami evolutivi corrispondenti, ovviamente, si fusero. Questa visione dell’evoluzione è chiamata teoria della simbiogenesi.
Ora la teoria della simbiogenesi è generalmente accettata. È stato confermato con la stessa precisione con cui può essere confermata qualsiasi teoria riguardante l’evoluzione su larga scala. Ma i concetti scientifici, a differenza dei dogmi religiosi, non rimangono mai statici. Naturalmente, il quadro generale della simbiogenesi oggi non ci sembra proprio lo stesso (e in alcuni luoghi non è affatto lo stesso) come lo immaginava Lynn Margulis mezzo secolo fa.
Logica classica
Nel cinquantesimo anniversario della pubblicazione del celebre articolo sulla simbiogenesi Giornale di biologia teorica ha preparato un numero speciale interamente dedicato all'eredità creativa di Lynn Margulis. Questo numero include un articolo completo del famoso biochimico e divulgatore scientifico britannico Nick Lane, che confronta lo stato attuale della questione dell'origine degli eucarioti con le idee classiche sull'argomento. Lane non ha dubbi che Margulis avesse ragione nelle affermazioni principali (riguardanti l'origine dei mitocondri e dei cloroplasti); ai nostri giorni, a quanto pare, nessuno degli scienziati seri ne dubita, perché i dati della biologia molecolare su questo argomento non sono ambigui. Ma il diavolo, si sa, vive nei dettagli. In questo caso, immergendoci nei dettagli, possiamo trovare molte cose nuove e interessanti e, soprattutto, assicurarci che l'argomento dell'origine degli eucarioti sia tutt'altro che esaurito.
Cominciamo dal fatto che alcune ipotesi private di Margulis si sono rivelate errate. Questo è normale: data l'enorme velocità di sviluppo della biologia, è semplicemente incredibile che assolutamente tutto sia stato indovinato con precisione in un articolo pubblicato mezzo secolo fa. Nuovi fatti che all'epoca non potevano essere conosciuti dall'autore apporteranno sicuramente alcuni aggiustamenti. Questo è quello che è successo anche qui. Innanzitutto Margulis insisteva sull’origine simbiotica non solo dei mitocondri e dei cloroplasti, ma anche dei flagelli eucariotici. Credeva che gli antenati dei flagelli fossero batteri mobili lunghi, attorcigliati a spirale, attaccati a una cellula eucariotica, simili alle moderne spirochete (vedi Fig. 1). Purtroppo questa ipotesi non ha ricevuto alcuna conferma biologica molecolare e ora nessuno la supporta più.
In alcuni momenti, Margulis avrebbe potuto avere ragione (questo non è proibito né dalle leggi della natura né dalla logica interna della sua stessa teoria), ma tuttavia, per ragioni indipendenti dalla sua volontà, ha sbagliato. Ad esempio, credeva che poiché i mitocondri discendono dai batteri, prima o poi i biologi impareranno a coltivarli in un mezzo nutritivo al di fuori delle cellule eucariotiche - beh, come i normali microbi. Se ciò fosse possibile, sarebbe la prova ideale della teoria della simbiogenesi. Purtroppo, infatti, i mitocondri moderni sono fondamentalmente incapaci di sopravvivenza indipendente, perché la maggior parte dei loro geni, durante l'evoluzione, sono migrati nel nucleo cellulare e lì sono stati integrati nel genoma dell '"ospite" eucariotico. Ora i prodotti proteici di questi geni vengono sintetizzati all'esterno del mitocondrio e quindi trasportati al suo interno utilizzando speciali sistemi di trasporto appartenenti alla cellula eucariotica. I geni rimasti nei mitocondri stessi sono sempre pochi: non sono sufficienti per sostenere la vita. Nel 1967 ancora nessuno lo sapeva.
Tuttavia, nel complesso, tutto ciò è particolare. Il pensiero di Lynn Margulis era sintetico: non si limitava a spiegare fatti individuali, ma cercava di combinarli in un sistema integrale che descrivesse l'evoluzione degli organismi viventi nel contesto della storia della Terra (Fig. 2). La moderna conoscenza scientifica consente di testare la forza di questo sistema di idee.

Albero e rete
Tutto è iniziato con l'ossigeno. Non c'era ossigeno molecolare (O 2) nell'antica atmosfera terrestre. Quindi i cianobatteri, che furono i primi a padroneggiare la fotosintesi dell'ossigeno, iniziarono a rilasciare questo gas nell'atmosfera (per loro era semplicemente un sottoprodotto non necessario). Nel frattempo, l'ossigeno puro è una sostanza molto tossica per chiunque non disponga di speciali mezzi biochimici di protezione contro di esso. Non sorprende che le emissioni di ossigeno dei cianobatteri abbiano avvelenato l’atmosfera terrestre e portato a un’estinzione di massa. Cominciò l’”olocausto dell’ossigeno” (L. Margulis, D. Sagan, 1997. Microcosmos: quattro miliardi di anni di evoluzione microbica).
Qui è già necessario un emendamento. Molti ricercatori moderni ritengono che la transizione da una biosfera priva di ossigeno a una biosfera basata sull’ossigeno sia stata in realtà molto più graduale e meno distruttiva di quanto suggerisca la speculazione sull’”olocausto dell’ossigeno” (vedi, ad esempio: “Il grande evento dell’ossigeno” a cavallo tra dell'Archeano e del Proterozoico non è stato grandioso, non è stato un evento, “Elements”, 03/02/2014). Inoltre, è possibile che la comparsa di ossigeno libero sia ancora più probabile è aumentato diversità di microrganismi, perché l’ossidazione di un certo numero di minerali da parte dell’ossigeno atmosferico ha arricchito la composizione chimica dell’ambiente e creato nuove nicchie ecologiche (M. Mentel, W. Martin, 2008. Energy Metabolism between eukaryotic anaerobes in light of Proterozoic ocean chemistry ). In generale, l'idea della comparsa dell'ossigeno nell'atmosfera come una grandiosa catastrofe che ha diviso l'intera storia della Terra in “prima” e “dopo” sembra ormai superata.
In un modo o nell'altro, non c'è dubbio che gli alfa-proteobatteri hanno beneficiato maggiormente dell'arricchimento del nostro pianeta con l'ossigeno. Hanno imparato a utilizzare direttamente l'ossigeno per produrre energia, e con grande efficienza. Ma gli antenati unicellulari degli eucarioti non avevano tale capacità. Erano anaerobici, cioè non potevano respirare ossigeno. Ma erano predatori che impararono ad assorbire cellule più piccole attraverso la fagocitosi. E questo ha dato loro un’ottima opportunità: catturare alcuni batteri, non digerendoli, ma “schiavizzandoli” e appropriandosi dei prodotti del loro metabolismo. Dopo aver assorbito l'alfa-proteobatterio, gli eucarioti primitivi erano in grado di respirare ossigeno: è così che si sono formati i mitocondri. E dopo aver assorbito il cianobatterio, è stato in grado di fotosintetizzare: è così che si sono formati i cloroplasti. Margulis riteneva che tali eventi si fossero verificati molte volte, seguendo una tendenza generale emersa. Questa è la cosiddetta sceneggiatura endosimbiosi seriale.
Quindi, Margulis scopre che a un certo stadio dello sviluppo della vita, l'endosimbiosi è diventata quasi un modello universale. Quindi, alla base dell'albero evolutivo degli eucarioti dovrebbe esserci letteralmente un'intera rete di rami evolutivi, che si intersecano tra loro a causa di eventi endosimbiotici e "crescono" approssimativamente nella stessa direzione - in quella dettata dalla combinazione di le allora condizioni esterne con le caratteristiche strutturali delle cellule (Fig. 3, A ).
Va detto che alla fine del XX secolo nella biologia evoluzionistica (e soprattutto nella paleontologia), l'idea che la maggior parte dei principali eventi evolutivi abbiano un carattere naturale e sistemico aveva già guadagnato una certa popolarità. Un tale evento copre contemporaneamente molti rami evolutivi, in cui, sotto l'influenza dell'eredità comune, sorgono approssimativamente le stesse caratteristiche in parallelo (vedi, ad esempio: A. G. Ponomarenko, 2004. Artropodizzazione e le sue conseguenze ecologiche). Esempi di tali eventi erano chiamati mammiferizzazione (l'origine dei mammiferi), angiospermizzazione (l'origine delle piante da fiore), artropodizzazione (l'origine degli artropodi), tetrapodizzazione (l'origine dei vertebrati terrestri), ornitizzazione (l'origine degli uccelli) e molto altro ancora. . Sembrava che la formazione degli eucarioti - l'eucariotismo - si inserisse perfettamente in questa serie.
Ad esempio, Kirill Eskov nel suo meraviglioso libro “La storia della Terra e la vita su di essa” (scritto negli anni '90) afferma quanto segue: “Molto probabilmente, diverse varianti di eucarioticità, cioè colonie intracellulari, sono comparse molte volte (per Ad esempio, c'è motivo di credere che le alghe rosse, che differiscono nettamente da tutte le altre piante in molte caratteristiche chiave, siano il risultato di tale "eucariotizzazione indipendente" dei cianobatteri)” (K. Yu. Eskov, 2000. History of the Earth and vita su di esso).
Purtroppo, in relazione agli eucarioti (non stiamo discutendo di altri esempi di “-azioni” ora), i dati moderni mettono in dubbio questo bellissimo scenario.
Problema dei mitocondri
Partiamo dal fatto che l'ipotesi sulle alghe rosse discussa da Eskov è ormai superata. Studi molecolari mostrano che il lignaggio evolutivo delle alghe rosse si trova nelle profondità dell'albero eucariotico (sono parenti abbastanza stretti delle piante verdi) e la loro eucarioticizzazione indipendente è estremamente improbabile.
Ma qualcos'altro è molto più serio. Se la simbiogenesi fosse un processo naturale, lungo, a più fasi, e procedesse addirittura in parallelo in diversi rami evolutivi, allora ci aspetteremmo di vedere uno spettro di stati di transizione piuttosto diversi tra eucarioti e non eucarioti. Questo è esattamente ciò che pensava Margulis. Il fatto che questi stati di transizione non siano evidenti, lei (per quanto si può giudicare) considerava un problema puramente tecnico associato alla mancanza di conoscenza e metodi imperfetti. È vero questo ora che sappiamo infinitamente di più sulle cellule viventi di quanto ne sapessimo cinquant’anni fa?
Ipotizziamo. Si supponeva che la presunta endosimbiosi seriale dovesse procedere, in primo luogo, gradualmente e, in secondo luogo, in modo leggermente diverso nelle diverse linee evolutive (poiché non ci sono ripetizioni esatte nell'evoluzione). Sulla base di ciò, Margulis predisse che prima o poi sarebbero stati scoperti eucarioti che avevano cloroplasti, ma non avevano mai mitocondri; eucarioti che hanno conservato flagelli batterici (che differiscono nettamente nella struttura dai flagelli degli eucarioti); e infine, principalmente eucarioti anaerobici, nelle cui cellule non ci sono tracce di adattamento all'atmosfera di ossigeno. Nessuna di queste previsioni è stata confermata. Nessuno degli eucarioti ha nemmeno un accenno di flagelli di tipo batterico: i loro mezzi di movimento sono completamente diversi. Nessuno degli eucarioti conosciuti può essere definito anaerobio primario: tutti, senza eccezioni, ad un certo punto della loro evoluzione hanno attraversato la "fase dell'ossigeno". Infine, tutti gli eucarioti hanno mitocondri attivi o i loro resti che hanno perso una parte significativa delle loro funzioni (idrogenosomi, mitosomi) o, nel peggiore dei casi, geni mitocondriali che sono riusciti a spostarsi nel nucleo.
Alla fine del 20° secolo, era diffusa l'ipotesi che alcuni moderni eucarioti unicellulari non avessero e non abbiano mai avuto mitocondri. È stato proposto di allocare tali eucarioti principalmente non mitocondriali in un regno speciale Archezoa. Margulis accettò questa ipotesi abbastanza presto e le rimase fedele fino all'ultimo, anche quando molti altri scienziati l'avevano già rifiutata (L. Margulis et al., 2005. “Imperfezioni e stranezze” nell'origine del nucleo). Riteneva molto probabile che gli eucarioti principalmente non mitocondriali (“archeoprotisti”) vivano ancora in alcuni habitat inaccessibili privi di ossigeno, dove sono molto difficili da individuare. Purtroppo, non sono stati ancora trovati "archeproti", ma sono stati trovati numerosi resti di mitocondri in quegli organismi unicellulari precedentemente classificati come Archezoi. Al momento si conosce un solo eucariota che non presenta alcuna traccia di mitocondri: il flagellato Monocercomonoidi, ma la posizione di questa creatura sull'albero evolutivo non lascia dubbi sul fatto che un tempo avesse mitocondri (A. Karnkowska et al., 2016. Un eucariota senza organello mitocondriale). In generale, al momento, senza eccezioni, tutti i casi di assenza di mitocondri negli eucarioti devono essere considerati secondari. Ciò significa che non esisteva uno stadio antico non mitocondriale nella storia degli eucarioti - almeno nei loro gruppi moderni -.
Margulis credeva (abbastanza ragionevolmente per il suo tempo) che in un certo periodo della storia della vita, l'eucariotismo fosse una tendenza ampia - una "tendenza", come si dice adesso. Sulla base di ciò, si potrebbe supporre che diversi eucarioti abbiano antenati diversi: ad esempio, che le alghe eucariotiche si siano evolute dai cianobatteri, gli animali da batteri predatori e i funghi da batteri osmotrofi che assorbono i nutrienti attraverso la superficie cellulare. Questa ipotesi non contraddice alcuna legge fondamentale della biologia. Ma, sfortunatamente, contraddice sorprendentemente i fatti. La sistematica molecolare mostra che l'antenato comune di piante, animali e funghi non era una forma transitoria, ma un vero eucariota, "a tutti gli effetti", come dice Nick Lane. Possiamo tranquillamente affermare che l'antenato comune di tutti gli eucarioti moderni era già una cellula eucariotica a tutti gli effetti: aveva un nucleo, un reticolo endoplasmatico, un apparato di Golgi, microtubuli, microfilamenti, mitocondri e flagelli. In generale, un insieme completo di caratteristiche eucariotiche.
Si prega di notare che questo insieme di caratteristiche non include i cloroplasti. Non sono comparsi in tutti gli eucarioti e non immediatamente. Inoltre, i cloroplasti furono certamente acquisiti più di una volta e in modi diversi in diversi rami evolutivi. I cloroplasti sono simili primario(quando un eucariote invade un cianobatterio), e secondario(quando un eucariota cattura un altro eucariote con un cianobatterio al suo interno) e persino terziario(quando un eucariota cattura un secondo eucariote, all'interno del quale vive un terzo eucariote, e all'interno di quello - un cianobatterio). Qui l'evoluzione, come si suol dire, è in pieno svolgimento. Con i mitocondri la situazione è completamente diversa: in base alla loro presenza non vediamo alcuna diversità particolare e nessuna fase transitoria (ad eccezione di numerosi fatti di perdita secondaria, ma tali fatti non dicono assolutamente nulla sull'origine degli eucarioti). Se lo scenario di Margulis fosse completamente corretto, la situazione con mitocondri e flagelli sarebbe più o meno la stessa di quella con i cloroplasti, ma non è così.
Ciò su cui Margulis aveva ragione era che gli eucarioti in generale sono piuttosto predisposti a prendere il sopravvento sugli endosimbionti. Qui possiamo fornire una serie di esempi, inclusa l'acquisizione di simbionti batterici da parte di alcuni vermi di acque profonde, sui quali questi vermi vivono effettivamente (V.V. Malakhov, 1997. I Vestimentiferi sono animali autotrofi). La rapida evoluzione dei cloroplasti è la manifestazione più sorprendente di questa tendenza. Solo gli “attori” che li acquisirono apparentemente possedevano già a quel tempo un insieme completo di caratteristiche eucariotiche, compresi i mitocondri. La configurazione dell'albero evolutivo degli eucarioti, come lo conosciamo oggi, semplicemente non consente altre versioni.
A ciò, Lane aggiunge che la struttura di base delle cellule differisce sorprendentemente poco tra i diversi eucarioti, a seconda del loro stile di vita (anche se lo stile di vita stesso può variare notevolmente). Tutti i componenti caratteristici di una cellula che la rendono eucariotica sono generalmente disposti allo stesso modo nelle piante, negli animali, nei funghi, nei flagellati e nelle amebe... "Ora sappiamo che quasi tutte le differenze tra gli eucarioti riflettono adattamenti secondari." , scrive Lane nell'articolo in discussione. L'uniformità della struttura della cellula eucariotica fa sì che i primi stadi della sua formazione non abbiano lasciato praticamente tracce nella moderna diversità degli eucarioti.
Evento unico
Le conclusioni che Lane trae oggi non possono più essere definite nuove o inaspettate. I dati moderni sono più compatibili con il presupposto che la formazione della cellula eucariotica sia avvenuta singolo evento, completato (nei tempi a nostra disposizione) molto rapidamente. È probabile che in questa fase gli antenati degli eucarioti attraversarono una sorta di collo di bottiglia (in un articolo precedente, Lane suggerì che si trattasse di una popolazione piccola, instabile e di breve durata nella quale ebbero luogo tutti i maggiori cambiamenti; N. Lane, 2011. Energetica e genetica attraverso il divario procarioti-eucarioti). Di conseguenza, nacquero i primi eucarioti "a pieno titolo", i cui discendenti si dispersero in diverse nicchie ecologiche, ma la struttura fondamentale della cellula non cambiò più. Pertanto, non vi era alcuna eucariotizzazione parallela. In ogni caso, la biologia moderna non trova prove che lo confermino.
I dati della genomica comparativa suggeriscono che l'evento soglia che separava gli eucarioti dal resto della natura vivente era l'unione di due cellule: una archaea (probabilmente appartenente a uno dei Lokiarchaeota) e una batterica (probabilmente appartenente a uno dei Proteobacteria) . Il superorganismo risultante divenne il primo eucariote (Fig. 3, B). Il moderno punto di vista “mainstream” identifica questo evento con l’acquisizione dei mitocondri (il cosiddetto scenario “early mitochondrial”; vedi, ad esempio: N. Yutin et al., 2009. The origins of phagocytosis and eukaryogenesis). In effetti, i mitocondri sono indiscutibili discendenti dei proteobatteri, e certamente penetrarono come simbionti nella cellula di un archaea (o di un eucariota primitivo non troppo lontano dagli archaea). Tuttavia, alla domanda su come siano arrivati lì, Lane dà una risposta piuttosto inaspettata. Vale a dire: “Non lo sappiamo”.
Qual è il problema? Secondo la teoria classica, tutti i simbionti interni venivano acquisiti dalle cellule eucariotiche mediante fagocitosi, cioè cattura da parte di pseudopodi con isolamento dell'oggetto catturato e successiva digestione (in questo caso fallita). Ciò è apparentemente vero per i cloroplasti, ma molto dubbio per i mitocondri. L'ipotesi che la fagocitosi sia comparsa prima dei mitocondri non si adatta bene ai dati bioinformatici. Un'analisi comparativa delle sequenze proteiche mostra che i microfilamenti di actina che formano la struttura interna di qualsiasi pseudopodo erano molto probabilmente inizialmente immobili - le proteine che consentono loro anche di contrarsi sono apparse molto più tardi (E.V. Kunin, 2014. Logica del caso). Ciò significa che l'evoluzione degli eucarioti non potrebbe iniziare direttamente con la fagocitosi: i mitocondri sono stati acquisiti in qualche altro modo.
Ma va sottolineato che tutto ciò è ancora solo speculazione. Il mistero dell'origine dei mitocondri, per non parlare dell'origine del nucleo, non è ancora stato risolto.
Possibilità e necessità
Quindi, l’ipotesi dell’endosimbiosi seriale è corretta? Sì, nel senso che gli eventi simbiotici si sono verificati molte volte nella storia degli eucarioti. Ciò è meglio illustrato dalla lunga, ricca e ormai ben studiata storia dei cloroplasti (P. Keeling et al., 2013. The number, speed, and Impact of plastid endosimbiosi in eukaryotic Evolution). No, nel senso che l'endosimbiosi seriale non era un prerequisito per l'emergere degli eucarioti come gruppo. L'evento endosimbiotico che portò alla comparsa degli eucarioti fu, per quanto possiamo giudicare oggi, unico.
Pertanto, lo scenario di “eucariotizzazione parallela” non è confermato. Ciò non significa che eventi evolutivi di questo tipo non accadano affatto: alcuni di essi sono descritti dettagliatamente dai paleontologi (ad esempio, la mammiferizzazione di rettili di tipo animale, che acquisiscono le caratteristiche dei mammiferi parallelamente in più rami evolutivi) . Inoltre, l’elenco di tali “scenari paralleli” è addirittura cresciuto di recente. "Elements" ha scritto più di una volta sull'ipotesi dell'emergere indipendente del sistema nervoso in due rami completamente diversi di animali multicellulari (vedi La discussione sul ruolo dei ctenofori nell'evoluzione continua, "Elements", 18.09.2015). Ma l’emergere degli eucarioti è uno degli eventi più singolari nell’intera storia della vita sulla Terra. Questo è probabilmente il motivo per cui cade da questa serie.
Nella letteratura scientifica moderna esiste un concetto come ipotesi delle terre rare(vedi ipotesi delle terre rare). I sostenitori di questa ipotesi ammettono che la vita relativamente semplice (a livello di organizzazione batterica) può esistere su molti pianeti ed essere abbastanza comune nell'Universo. Ma la vita relativamente complessa (eucariotica o paragonabile) nasce solo nella più rara combinazione di circostanze; è possibile che esista un solo pianeta con tale vita nella Galassia. Se l’ipotesi della Terra rara è corretta, allora l’emergere degli eucarioti è molto probabilmente l’evento fondamentale che separa la vita “semplice” (diffusa) dalla vita “complessa” (improbabile).
L'autore del famoso libro "L'origine della vita", Mikhail Nikitin, è giunto recentemente (e in modo completamente indipendente) a conclusioni simili. “Non sappiamo ancora quanto sia stata naturale la comparsa degli eucarioti. Se per altre fasi dello sviluppo della vita, come la transizione dal mondo dell'RNA al mondo delle proteine dell'RNA, la separazione delle cellule procariotiche dal "mondo dei virus" precellulare o l'emergere della fotosintesi, possiamo dire con sicurezza che sono naturali e quasi inevitabili, poiché la vita è già apparsa, quindi la comparsa degli eucarioti nella biosfera procariotica potrebbe essere molto improbabile. È possibile che nella nostra Galassia ci siano miliardi di pianeti con vita a livello batterico, ma solo sulla Terra sono comparsi gli eucarioti, sulla base dei quali sono comparsi animali multicellulari e poi esseri intelligenti” (M. Nikitin, 2014. Una nuova ipotesi dell'origine della cellula eucariotica). Forse è per questo che ci risulta così difficile comprendere nei dettagli l'origine degli eucarioti: si tratta di un evento unico (su scala planetaria), al quale è molto difficile applicare il principio dell'uniformitarismo, che richiede “per impostazione predefinita” ” procedere dall’uniformità di fattori e processi in ogni momento. Ma proprio per questo il mistero dell'origine degli eucarioti è uno dei più affascinanti di tutta la biologia. Ci sono ancora molte questioni irrisolte in questo ambito; non tutte sono menzionate qui (come nell'articolo discusso da Nick Lane).
Ministero della Sanità dell'Ucraina
Università medica statale di Zaporozhye
Dipartimento di Microbiologia, Virologia e Immunologia
Abstract sull'argomento:
"Teorie sull'origine della rickettsia e dei mitocondri"
preparato da uno studente
3 portate 30 gruppi
Mikheeva Evgenia Sergeevna
Zaporozhye
1. Teoria della simbiogenesi
2. Rickettsia
3. Scoperto il parente più stretto dei mitocondri
4. Nick Lane
5. Estratti dal popolare libro scientifico di Nick Lane “Energia, sesso, suicidio. I mitocondri e il significato della vita"
6. Caratteristiche del metabolismo
Bibliografia
Teoria della simbiogenesi
(teoria simbiotica, teoria endosimbiotica, teoria dell'endosimbiosi) spiega il meccanismo di comparsa di alcuni organelli delle cellule eucariotiche: mitocondri, idrogenosomi e plastidi.
La teoria dell'origine endosimbiotica dei cloroplasti fu proposta per la prima volta nel 1883 Andrea Schimper, che hanno mostrato la loro autoreplicazione all'interno della cellula. La sua apparizione è stata preceduta dalla conclusione A. S. Famintsina e O.V. Baranetsky sulla duplice natura dei licheni del complesso simbiotico di funghi e alghe (1867). K. S. Merezhkovsky nel 1905 propose il nome stesso "simbiogenesi", per la prima volta formulò la teoria in dettaglio e creò persino un nuovo sistema del mondo organico sulla sua base. Anche Famintzin nel 1907, basandosi sul lavoro di Schimper, arrivò alla conclusione che i cloroplasti sono simbionti, come le alghe nei licheni.
Negli anni '20, la teoria fu sviluppata da B. M. Kozo-Polyansky e fu suggerito che anche i mitocondri fossero simbionti. Quindi, per molto tempo, la simbiogenesi non è stata praticamente menzionata nella letteratura scientifica. La teoria ampliata e concretizzata ha ricevuto una rinascita nei lavori di Lynn Margulis a partire dagli anni '60.
Origine simbiotica dei mitocondri e dei plastidi
Come risultato dello studio della sequenza delle basi nel DNA mitocondriale, sono stati ottenuti argomenti molto convincenti a favore del fatto che i mitocondri sono i discendenti di batteri aerobi (procarioti) legati alla rickettsia, che una volta si stabilirono nella cellula eucariotica ancestrale e "impararono" viverci come simbionti. Ora i mitocondri sono presenti in quasi tutte le cellule eucariotiche; non sono più in grado di riprodursi al di fuori della cellula.
Esistono prove che gli antenati endosimbiotici originali dei mitocondri non potevano né importare proteine né esportare ATP. Probabilmente inizialmente ricevettero piruvato dalla cellula ospite e il beneficio per l'ospite fu la neutralizzazione dell'ossigeno tossico per il nucleocitoplasma da parte dei simbionti aerobici.
La teoria simbiotica dell'origine degli organelli è diventata un classico della biologia moderna. All'ordine del giorno ci sono le questioni della sua concretizzazione, costruzione della filogenesi, simbiosi, ricerca di legami familiari, in generale, un lavoro ampio e importante, senza il quale è impossibile “costruire un ponte” dalla teoria alla pratica. Numerosi dati indicano che l'origine tassonomica dei mitocondri è nell'ordine Rickettsiales. Ricordiamo che le rickettsie, come i mitocondri, non possono esistere al di fuori della cellula ospite, ma a differenza di quest'ultima, spesso danneggiano l'ospite, provocando ad esempio il tifo. Il nostro connazionale V.V. Emelyanov dell'Istituto di Epidemiologia e Microbiologia da cui prende il nome. NF Gamaleya e i suoi colleghi nel 2001 proposero l'ipotesi che la moderna rickettsia patogena e i mitocondri avessero un antenato comune, simile agli endosimbioni simili a rickettsia (RPE) che coesistono con i parameci. Secondo gli autori dell'opera (V.V. Emelyanov, M.Yu. Vysokikh), "l'ultimo antenato comune non solo ha perso i geni ridondanti..., ma ha anche trasferito alcuni geni vitali nel genoma ospite". Utilizzando l'immunoblotting di proteine provenienti da cellule intere e frazioni di membrana di Rickettsia prowazekii (l'agente eziologico del tifo), si è scoperto che una delle proteine codificate da tali geni è la proteina della membrana esterna dei mitocondri - porina, che è codificata dal nucleo, importato nei mitocondri, ma viene utilizzato anche, funzionalmente, da Rickettsia prowazekii. Questa osservazione è, se non una prova, una conferma molto seria dell'ipotesi.
Sebbene le rickettsie siano di dimensioni paragonabili ad alcuni virus, sono chiaramente distinte da questi. Le cellule di Rickettsia contengono sia DNA che RNA (in un rapporto di 1:3,5); sono circondati da una parete cellulare contenente acido muramico e sensibile al lisozima. Le micrografie elettroniche di sezioni ultrasottili mostrano la regione nucleare e la parete cellulare.
La maggior parte delle rickettsie non sono mai state coltivate al di fuori di una cellula vivente, ma possono propagarsi nelle uova incubate e nei tessuti animali; Dal sacco vitellino di un uovo di gallina puoi ottenere 10 9 cellule. Nelle cellule isolate di rickettsia possono essere rilevati alcuni enzimi metabolici intermedi. Durante la coltivazione, il tasso metabolico di tali cellule si indebolisce, ma l'aggiunta di ATP, acidi organici e aminoacidi stimola nuovamente la loro respirazione. Le Rickettsie hanno quindi un proprio metabolismo; tuttavia, probabilmente a causa di cambiamenti nella permeabilità della superficie cellulare, non sono in grado di regolare l'assorbimento e l'escrezione dei metaboliti.
Informazioni correlate.
La teoria della simbiogenesi (teoria simbiotica, teoria endosimbiotica o teoria endosimbiosi) spiega il meccanismo dell'emergere di alcuni organelli della cellula eucariotica: mitocondri, plastidi e idrogenosomi.
L'essenza del concetto è la coesistenza reciprocamente vantaggiosa di un organello con una cellula. Ciò suggerisce l'endosimbiosi come una simbiosi vantaggiosa per entrambe le parti con la formazione di cellule eucariotiche (cellule in cui è presente un nucleo). Quindi, utilizzando il trasferimento di informazioni genetiche tra batteri, sono stati effettuati il loro sviluppo e l'aumento della popolazione. Secondo questa versione, ogni ulteriore sviluppo della vita e delle forme di vita è dovuto al precedente antenato delle specie moderne.La teoria dell'origine endosimbiotica dei cloroplasti fu proposta per la prima volta nel 1883 da Andreas Schimper, che dimostrò la loro autoreplicazione all'interno della cellula.
La sua comparsa fu preceduta dalla conclusione di A. S. Famintsyn e O. V. Baranetsky sulla duplice natura dei licheni: un complesso simbiotico di funghi e alghe (1867).
Le chiare disposizioni del sistema furono redatte dal botanico e zoologo russo K. S. Merezhkovsky.
Nel 1905 propose il nome stesso "simbiogenesi", per la prima volta formulò la teoria in dettaglio e creò sulla base un nuovo sistema del mondo organico. Anche Famintsyn nel 1907, basandosi sul lavoro di Schimper, giunse alla conclusione che i cloroplasti sono simbionti, come le alghe nei licheni.
Negli anni '20, la teoria fu sviluppata da B. M. Kozo-Polyansky e fu suggerito che anche i mitocondri fossero simbionti. Quindi, per molto tempo, la simbiogenesi non è stata praticamente menzionata nella letteratura scientifica. La teoria ampliata e concretizzata ha ricevuto una rinascita nei lavori di Lynn Margulis, a partire dagli anni '60.
Come risultato dello studio della sequenza delle basi nel DNA mitocondriale, sono stati ottenuti argomenti molto convincenti a favore del fatto che i mitocondri sono i discendenti di batteri aerobi (procarioti) legati alla rickettsia, che una volta si stabilirono nella cellula eucariotica ancestrale e "impararono" viverci come simbionti. Ora i mitocondri sono presenti in quasi tutte le cellule eucariotiche; non sono più in grado di riprodursi al di fuori della cellula.
Esistono prove che gli antenati endosimbiotici originali dei mitocondri non potevano né importare proteine né esportare ATP. Probabilmente inizialmente ricevettero piruvato dalla cellula ospite; il beneficio per l'ospite era la neutralizzazione dell'ossigeno tossico per il nucleocitoplasma da parte dei simbionti aerobici.
I plastidi, come i mitocondri, hanno il proprio DNA procariotico e ribosomi. Apparentemente, i cloroplasti provenivano da batteri fotosintetici, che un tempo si stabilirono nelle cellule eterotrofe dei protisti e le trasformarono in alghe autotrofe.
Prova
Mitocondri e plastidi:
Hanno due membrane completamente chiuse. In questo caso quella esterna è simile alle membrane dei vacuoli, quella interna è simile ai batteri,
Si riproducono per fissione binaria (e talvolta si dividono indipendentemente dalla divisione cellulare), non vengono mai sintetizzati de novo,
Materiale genetico - DNA circolare non associato agli istoni (in termini di proporzione di GC, il DNA dei mitocondri e dei plastidi è più vicino al DNA dei batteri che al DNA nucleare degli eucarioti),
Possiedono un proprio apparato di sintesi proteica - ribosomi e altri ribosomi di tipo procariotico - con una costante di sedimentazione di 70S. La struttura dell'rRNA 16s è simile a quella dei batteri.
Alcune proteine di questi organelli sono simili nella loro struttura primaria a proteine simili dei batteri e non sono simili alle corrispondenti proteine del citoplasma.
I problemi
Il DNA dei mitocondri e dei plastidi, a differenza del DNA della maggior parte dei procarioti, contiene introni.
Solo una parte delle loro proteine sono codificate nel DNA dei mitocondri e dei cloroplasti, mentre il resto è codificato nel DNA del nucleo cellulare. Durante l'evoluzione, parte del materiale genetico “fluisce” dal genoma dei mitocondri e dei cloroplasti al genoma nucleare. Ciò spiega il fatto che né i cloroplasti né i mitocondri non possono più esistere (riprodursi) in modo indipendente.
La questione dell'origine della componente nucleare-citoplasmatica (NCC), che ha catturato i protomitocondri, non è stata risolta. Né i batteri né gli archei sono capaci di fagocitosi, nutrendosi esclusivamente per via osmotrofica. Studi biologici e biochimici molecolari indicano la natura chimerica arcaico-batterica della JCC. Anche come sia avvenuta la fusione di organismi provenienti da due domini non è chiaro.
Esempi di endosimbiosi
Al giorno d'oggi, esistono numerosi organismi che contengono altre cellule all'interno delle loro cellule come endosimbionti. Essi, tuttavia, non sono gli eucarioti primari sopravvissuti fino ai giorni nostri, nei quali i simbionti non si sono ancora integrati in un unico insieme e non hanno perso la loro individualità. Tuttavia, dimostrano in modo chiaro e convincente la possibilità della simbiogenesi.
Mixotricha paradoxa è l'organismo più interessante da questo punto di vista. Per muoversi utilizza più di 250.000 batteri spirochete Treponema attaccati alla superficie della sua cellula. I mitocondri di questo organismo sono andati perduti per la seconda volta, ma all'interno della sua cellula si trovano batteri aerobici sferici che sostituiscono questi organelli.
Anche le amebe del genere Pelomyxa non contengono mitocondri e formano una simbiosi con i batteri.
I ciliati del genere Paramecium contengono costantemente alghe all'interno delle loro cellule, in particolare il Paramecium bursaria forma endosimbiosi con le alghe verdi del genere Chlorella.
L'alga flagellata unicellulare Cyanophora paradoxa contiene cianelli, organelli che assomigliano ai tipici cloroplasti delle alghe rosse, ma differiscono da essi per la presenza di una sottile parete cellulare contenente peptidoglicano. La dimensione del genoma della cianella è uguale a quella dei tipici cloroplasti e molte volte inferiore a quella dei cianobatteri.
Ipotesi sull'origine endosimbiotica di altri organelli
L'endosimbiosi è la versione più ampiamente accettata dell'origine dei mitocondri e dei plastidi. Ma i tentativi di spiegare l'origine di altri organelli e strutture cellulari in modo simile non trovano prove sufficienti e incontrano critiche giustificate.
Perossisomi
Christian de Duve scoprì i perossisomi nel 1965. Ha anche suggerito che i perossisomi fossero i primi endosimbionti di una cellula eucariotica, permettendole di sopravvivere con una quantità crescente di ossigeno molecolare libero nell’atmosfera terrestre. I perossisomi, però, a differenza dei mitocondri e dei plastidi, non hanno né materiale genetico né un apparato per la sintesi proteica. È stato dimostrato che questi organelli si formano de novo nella cellula del RE e non c'è motivo di considerarli endosimbionti.