Poreklo eukariota. Simbiotska teorija, endosimbioza ili simbiogeneza Simbiotska teorija
Treba napomenuti tri hipoteze o porijeklu eukariotskih ćelija:
- simbiotska hipoteza, ili simbiogeneza,
- intususcepcija,
- himerni.
Danas je u naučnom svijetu priznata glavna hipoteza porijekla eukariota simbiogeneza.
Prema simbiogenezi, organele eukariotskih stanica kao što su mitohondrije, hloroplasti i flagele nastale su uvođenjem nekih prokariota u drugu, veću prokariotsku ćeliju, koja je igrala ulogu ćelije domaćina.
Simbiotska hipoteza ima poteškoća u objašnjavanju porijekla jezgra eukariotskih ćelija i u pitanju koji je prokariot bio domaćin. Podaci iz molekularne analize genoma i proteina eukariota pokazuju da se, s jedne strane, radilo o organizmu bliskom arheji (ranije klasifikovane kao bakterije, a zatim su izdvojene u zasebnu granu). S druge strane, eukarioti sadrže proteine (i gene odgovorne za njihovu sintezu) koji su karakteristični za potpuno različite grupe prokariota.
Prema invaginacionoj hipotezi porekla eukariotskih ćelija, njihove organele su nastale invaginacijom citoplazmatske membrane sa naknadnim razdvajanjem ovih struktura. Formirano je nešto poput kuglica, okruženih membranom i koje sadrže unutar citoplazme i spojeve i strukture zarobljene ovdje. U zavisnosti od toga šta je uneseno, formirane su različite organele.
Prokarioti nemaju prave organele, njihove funkcije obavljaju invaginacije membrane. Stoga je lako zamisliti takvo razvezivanje. U prilog hipotezi invaginacije ide i sličnost citoplazmatske membrane i dvostrukih membrana organela.
Sa stajališta invaginogeneze, porijeklo jezgra je lako objasniti, ali je neobjašnjivo zašto se genom i ribosomi nuklearno-citoplazmatskog kompleksa razlikuju od onih u hloroplastima i mitohondrijima (zapamtite da imaju i DNK i ribozome). Štaviše, u ovim organelama, sistem biosinteze proteina (DNK, RNK, ribozomi) je sličan prokariotima.
Ova razlika je dobro objašnjena upravo sa stanovišta prve, simbiotske, hipoteze. Prema tome, anaerobni prokariot je nekako ušao u aerobni prokariot. Nije se probavio, već je, naprotiv, počeo da se hrani na račun ćelije domaćina. Zauzvrat je koristio kiseonik za dobijanje energije, a ovaj način oksidacije je mnogo efikasniji; dao je višak energije prokariotskom domaćinu, koji je u ovom slučaju takođe imao koristi. Nastala je simbioza. Nakon toga, invazivni prokariot se pojednostavio, dio njegovog genoma migrirao je u ćeliju domaćina i više nije mogao postojati samostalno.
Simbiogeneza na sličan način objašnjava porijeklo hloroplasta. Uvedeni su samo prokarioti sposobni za fotosintezu (slično plavo-zelenim algama).
Prvi eukarioti bez fotosintetskih simbionta iznjedrili su životinje u kojima su se pojavili - biljke.
Trenutno postoje protozoe (jednoćelijski eukariotski organizmi) koji nemaju mitohondrije ili hloroplaste. Ali umjesto njih, simbiontski prokarioti se naseljavaju u citoplazmi i obavljaju odgovarajuće funkcije. Ova činjenica, kao i sličnost sistema biosinteze proteina mitohondrija i plastida sa prokariotima, smatraju se dokazom simbiogeneze. Dokaz dolazi i iz činjenice da se mitohondrije i hloroplasti razmnožavaju nezavisno; ćelija ih nikada ne gradi od nule.
Treću, himernu, hipotezu o poreklu eukariotskih ćelija podržava velika veličina njihovog genoma, koja hiljadama ili više puta prevazilazi genom bakterije, kao i raznolikost sintetizovanih proteina koji se nalaze u različitim grupama prokariota. Jasno je da je tokom evolucije eukariota njihov genom postao složeniji, udvostručio se i u njemu su se pojavili mnogi regulatorni geni. Međutim, početno povećanje veličine genoma moglo se dogoditi zbog kombinacije genoma nekoliko prokariota.
Možda je u davna vremena određeni prokariot stekao sposobnost fagocitoze i, hraneći se na taj način, apsorbirao druge prokariote koji nisu uvijek bili probavljeni. Njihov genom je sadržavao gene korisne za domaćina, a on ih je uključio u svoj genom. Možda su neki od prokariota koji su završili unutra postali organele, što kombinuje himernu hipotezu sa simbiogenezom.
Budući da sve gore navedene hipoteze imaju prednosti i nedostatke, i na mnogo načina ne isključuju jedna drugu, onda je, po našem mišljenju, kombinacija mnogih faktora opisanih različitim hipotezama mogla igrati ulogu u nastanku struktura eukariotskih stanica. .
Također treba napomenuti da se prema simbiotičkoj hipotezi, porijeklo drugih membranskih organela kao što su vakuole, Golgijev kompleks itd. može smatrati daljnjim pojednostavljenjem, na primjer, mitohondrija.
Također primjećujemo da kombinacija proteina iz različitih prokariota u eukariotima možda uopće ne ukazuje na himerizam prvih. Moguće je da određene enzimske funkcije mogu obavljati samo proteini određene strukture. I evolucija eukariota je u više navrata dolazila do ove točke, bez obzira na evoluciju grupa prokariota koji nisu bili njihovi preci. Recimo da je došlo do konvergencije na molekularnom nivou.
Mitohondrije su vjerni pratioci eukariota. Prema teoriji simbiogeneze, upravo je stjecanje mitohondrija izazvalo stvaranje nuklearnih organizama. Jedan od dokaza ove teorije bilo je otkriće mitohondrija ili sličnih organela kod svih, čak i najjednostavnijih, eukariota. Ali u maju 2016. tim čeških naučnika opisao je prvi nuklearni organizam u istoriji koji nije sadržavao čak ni indirektne znakove mitohondrija. Može li ovo otkriće poljuljati današnje ideje o ranoj evoluciji eukariota?
Mitohondrije su dvomembranske organele koje opskrbljuju energijom stanice gotovo svih eukariota. Pouzdano je poznato da su u srodstvu sa α-proteobakterijama i da su postale dio eukariotskih ćelija prije oko 1,5 milijardi godina. Bakterijsko porijeklo mitohondrija dokazuje prisustvo dvije membrane (unutarnje vlastite i vanjske domaćine), vlastite kružne DNK i mašine za prevođenje, kao i sposobnost samostalne podjele. Neki čak vjeruju da je apoptoza pokušaj mitohondrija da ubije eksploatatora koji ju je apsorbirao.
Međutim, u naučnoj zajednici još uvijek ne postoji konsenzus o ulozi ove simbioze u razvoju eukariota (Sl. 1). Pristalice teorije simbiogeneze tvrde da je spajanje određene arheje s pretkom mitohondrija (bakterija) pokrenulo lanac događaja koji su doveli do formiranja modernih eukariota. Followers arheozojska teorija (hipoteza) Naprotiv, vjeruju da samo već formirani nuklearni organizam, arhezoan, može "ukrotiti" mitohondriju.
Zahvaljujući Carlu Woeseu i Georgeu Foxu, koji su 70-ih godina prošlog stoljeća upoređivali gene 16S rRNA mnogih živih bića, koji su svojom vizualnom sličnošću dugo obmanjivali mikrobiologe, dva oblika prenuklearnih organizama (prokariota) su potpuno i nepovratno razdvojena. , pa čak i na najvišem nivou: eubakterijama su oduzeta prava na “istinu” (-eu), arhebakterije imaju pravo da nose ponosno ime bakterija, ali su u novom sistemu živih organizama dobile svoj domen ( takson najvišeg ranga): “ Evolucija između kamena i nakovnja ili kako je mikrobiologija spasila evoluciju od apsorpcije molekularne biologije" i " Carl Woese (1928–2012)". Tako je 1990. godine od osobe zatraženo da shvati da su sva živa bića filogenetski podijeljena na tri domene: bakterije, arheje i eukariote, a bakterije se od arheja razlikuju čak više nego arheje od eukariota, a nedavno su gotovo posumnjali u preporučljivost podjele ovih potonjih. : " Pronađeni su preci svih eukariota". Međutim, prijedlog od prije skoro trideset godina još uvijek nije naišao na razumijevanje kod mnogih autora domaćih udžbenika biologije. I šta? Šta ako ti naučnici ponovo sve promene, pa svakih dvadesetak godina moraju da prepisuju udžbenike? - Ed.
Godine 1928., poznati biolog s prijelaza 19. u 20. vek, Edmund Wilson, govorio je o hipotezi o bakterijskom poreklu mitohondrija na sledeći način: “Takve ideje su previše fantastične da bi se o njima raspravljalo u pristojnom biološkom društvu.”. Danas se sličan stav formirao i prema teoriji arheozoika, te je općenito prepoznata ključna uloga mitohondrija u ranoj evoluciji eukariota. Otvaranje prvog pravi nemitohondrijski protozoan tjera vas da ponovo razmislite o prednostima i slabostima svake teorije.
Teorija simbiogeneze
Jedna od prednosti simbiogenetske teorije u odnosu na teoriju arheozoa je da objašnjava nastanak jezgra i intronsku strukturu genoma. Kod prokariota je široko rasprostranjen horizontalni transfer gena (HGT), zbog čega populacije mogu brzo razmjenjivati dijelove genskog fonda. HGT doprinosi nesigurnosti genoma - uostalom, ulazna DNK u ovom slučaju nije ni na koji način odvojena od sadržaja ćelije domaćina.
Vrlo vjerojatni pokušaji ćelije domaćina da uništi još neudomaćeni simbiont doveli su do oslobađanja simbiotske DNK u citoplazmu. Ova DNK, budući da je u neposrednoj blizini genoma domaćina, mogla bi se lako integrirati u njega. Zbog HGT, čak i kod eukariota koji su izgubili mitohondrije, u početku se nalaze mitohondrijski geni.
Takva fuzija genoma mogla bi, prije svega, doprinijeti razvoju međuzavisnosti između simbionta i domaćina. Drugo, obilni HGT mogao bi nositi ne samo gene koji su osiguravali preplitanje metabolizma dvaju organizama, već i sebične retroelemente. Invazija introna grupe II, koji su pobjegli od α-proteobakterije, dovela je do labavljenja prvobitno vrlo gustog genoma domaćina: do 80% DNK domaćina sada su introni. U tako složenoj situaciji, ćelija domaćin je razvila nekoliko linija odbrane svog genoma od salve introna: sistem unutrašnjih membrana i jezgra, ubikvitinski sistem za razgradnju oštećenih proteina, besmisleno posredovano raspadanje RNK i druge pojavile su se karakteristične osobine eukariota (slika 3).
Slika 3. Formiranje glavnih karakteristika eukariota može se objasniti invazijom introna grupe II u genom ćelije domaćina koja je uslijedila nakon mitohondrijalne simbioze.
Još jedan snažan dokaz za simbiogenezu su energetski zahtjevi eukariota. Iako je potrošnja energije proeukariota po gramu težine približno ista, ćelije s jezgrom su mnogo veće od ćelija bez jezgre, zbog čega troše otprilike 5000 puta više energije (2300 pW/ćeliji naspram 0,5 pW/ćeliji). Kada se preračuna potrošnja energije za prosječan gen jednoćelijskog organizma, ispada da eukariotski gen troši 1000 puta više energije. Bez mitohondrijske energije bilo bi nemoguće ne samo stvoriti složene, velike i aktivno pokretne organizme, već čak i osigurati funkcioniranje staničnih struktura tipičnih za eukariote.
Kod divovskih bakterija, skaliranje prokariotske energije zbog masovne poliploidizacije (kao u slučaju Epulopiscium, raste na 0,6 mm i sadrži 200.000 kopija genoma veličine 3,8 miliona bp) ne dovodi do povećanja izlazne energije po genu, a ćelija ostaje tipično bakterijska. - Auto.
Još jedna važna činjenica koja podržava simbiogenetski scenario je postojanje intracelularnih bakterijskih simbionta. Slučajevi endosimbioze kod bakterija su izuzetno rijetki u prirodi, ali i dalje postoje i pokazuju kako bi eukariotski domen života mogao nastati.
Archaeozoic theory
Arheozoi su navodni nemitohondrijski, ali nuklearni preci modernih eukariota. Prema arheozojskom scenariju, mitohondrije su pripitomljene tek u kasnim fazama eukariotske evolucije i nisu imale značajan uticaj na ovaj proces.
Jedna od glavnih odredbi simbiogeneze je hipoteza početne jednostavnosti. Vrlo malo se zna o životu tokom proterozoika, tako da postoje mnoge često međusobno isključive pretpostavke o njegovoj strukturi. Ako se, prema prvoj hipotezi, vjeruje da su složeniji eukarioti evoluirali iz prokariota s vrlo kompaktnim genomima, onda su u arheozojskom scenariju u početku postojale ćelije sa zbunjujućim i glomaznim genomima, iz kojih su jednostavniji prokarioti evoluirali redukcijom. Eukarioti su zadržali samo svoju primarnu složenost.
Evolucija genoma ne ide uvijek od jednostavnog ka složenom. A među eukariotima postoje primjeri koji to potvrđuju.
Ipak, smanjenje genoma ne mora nužno pratiti njegovo zbijanje. Dokazi za to mogu se naći i u protozojskim i višećelijskim oblicima života.
Na primjer, slobodno žive cilijate Paramecium tetraurelia sadrži 30.000 gena, od kojih svaki ima u prosjeku 2 kb. Ova kompaktnost se postiže smanjenjem veličine introna na maksimalnih 25 bp. i smanjenje međugenskih udaljenosti.
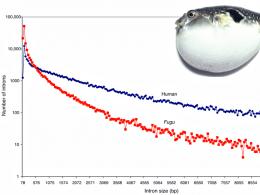
Čak i kralježnjaci mogu imati neobično kompaktne genome: genom ribe puf je osam puta manji od ljudi, uglavnom zbog malog sadržaja ponavljanja (slika 4).

Slika 4. Riba puhačica ima neobično kompaktan genom za kičmenjaka. dijelom zbog kratkih introna. Vertikalna os je data u logaritamskoj skali.

Slika 5. Hipoteza početne složenosti implicira da su prokariotske grane života evoluirale od složenijih oblika kroz redukciju. Vektor redukcije razvoja mogli su postaviti prvi grabežljivi arheozoi, ugnjetavajući druge organizme.
Gore navedeni primjeri pokazuju da jednostavnost prokariotskih genoma može nastati sekundarno. Ako je to istina, onda bi LUCA - posljednji zajednički predak svih modernih organizama - mogao imati eukariotski genom.
Hipotezu o početnoj složenosti potvrđuju i takozvani signaturni geni (“signature”) - eukariotski geni koji nemaju prokariotske homologe. Najvjerovatnije su ovi geni bili sadržani u LUCA, ali su ih bakterije i arheje izgubile.
Na nesreću arhezojske teorije, lista signatura se značajno prorijedila od početka 21. stoljeća. Među mnogim genomima sekvenciranim od tada, pronađeni su njihovi prokariotski homolozi. Tako je svake godine sve više proteina, čije se prisustvo kod eukariota može objasniti činjenicom da je njihove gene donio arhealni ili bakterijski predak tokom simbiogeneze.
A u isto vrijeme, otkriće prokariotskih homologa proteina staničnog kretanja (aktina, tubulina i kinezina) indirektno potvrđuje mogućnost da bi se arheozoi mogli aktivno kretati, pa čak i biti prvi grabežljivci na Zemlji sposobni za fagocitozu. Pojava grabežljivaca u autotrofno-saprotrofnoj zajednici kolijevke života trebala je imati kolosalan utjecaj na tok evolucije. U najuzbudljivijim scenarijima, neki arhezojski plijen prilagođavaju se brzoj podjeli i rastu, dok se drugi prilagođavaju nišama u koje arhezoan ne može prodrijeti. Kao rezultat toga, hipotetički arhezoan je vodio evoluciju svojih savremenika reduktivnim putem s naglaskom na metaboličkoj fleksibilnosti i stopi podjele, tokom koje su se formirali nama poznati prokarioti (slika 5).
Ali uprkos činjenici da arheozojska teorija ima neke jake tačke ili barem zadaje jake udarce simbiogenezi, nedostaje joj ono glavno - ne objašnjava kako i zašto je nastalo jezgro.
Jedinstveni nalaz
Osamdesetih godina prošlog stoljeća bilo je mnogo kandidata za titulu modernog arheozoa, ali narednih godina u svima su pronađene organele slične mitohondrijima (mitosomi i peroksizomi) i markerski geni mitohondrijske prošlosti: geni za sklapanje Fe- S proteini, mitohondrijski transporteri i šaperoni, kardiolipin sintetaza. Osim toga, neki proteini sintetizirani u citoplazmi imaju uvozne sekvence u mitohondrije, koje mogu opstati u odsustvu samih mitohondrija.
Sa svakim novim „zatvaranjem“ potencijalnog arheozoa, nemitohondrijski scenario za formiranje eukariota se pokazao sve manje verovatnim. A onda se u maju 2016. konačno pojavio novi potencijalni arhezoik, koji nije sadržavao ni trag mitohondrija.
Govorimo o anaerobnoj oksimonadi Monocercomonoides sp. PA203, koji živi u crijevima insekata. Oksimonade nemaju mitohondrije i ne sadrže gene mitohondrijalnog porijekla u svojoj nuklearnoj DNK. Oni primaju energiju od glikolize koja se odvija u citoplazmi.
Genom Monocercomonoides sp., koju je dešifrovao tim čeških naučnika, sadrži 16.629 gena, među kojima nema gore navedenih markera. Potraga za mitohondrijskim homolozima i proteinima sa uvoznim sekvencama takođe nije dala zadovoljavajuće rezultate (slika 6).
Jedino što je pronađeno su dva gena, čiji su proizvodi pronađeni kod bliskog rođaka Monocercomonoides sp. mogu (ili ne moraju) biti sadržane u mitohondrijama, ali im nedostaju uvozne sekvence.
U to vjeruju autori otkrića Monocercomonoides je nekada sadržavao mitohondrije, jer blisko srodni rodovi imaju tragove mitohondrija. Još uvijek je moguće da ove protozoe imaju još neotkrivene mitosome koji su postali toliko degradirani da nema dokaza o njihovom prisustvu u genomu.
u svakom slučaju, Monocercomonoides sp.- do sada jedinstven slučaj istinski bez mitohondrijalnog protista u čitavoj istoriji biologije. I ovaj slučaj dokazuje da eukarioti mogu živjeti ne samo bez mitohondrija, već i bez njihovog genetskog naslijeđa.
U kom su slijedu preci eukariota stekli unutarćelijske stvari i što je postalo sretna karta za evolucijsku budućnost, možete saznati iz članka " Genealogija proteina ukazuje na kasno sticanje mitohondrija od strane eukariotskih predaka» . - Ed.
Ovo otkriće, naravno, ne zadaje snažan udarac teoriji simbiogeneze, ali definitivno postavlja pitanja o tome šta je neophodno, a šta višak kod eukariota.
Književnost
- Kako su se pojavile mitohondrije (priča slična bajci);
- Kunin E.V. Logika slučaja. M.: Centrpoligraf, 2014. - 527 str.;
- Evolucija između kamena i nakovnja, ili Kako je mikrobiologija spasila evoluciju da je ne proguta molekularna biologija;
- Carl Woese (1928–2012);
- Kondratenko Y. (2015). "Pronašli smo pretke svih eukariota." "Shroedingerova mačka". 6 ;
- van der Giezen M. (2009).
Prije 50 godina, 1967., Lynn Margulis je objavila opširno izvješće o simbiogenetskoj teoriji, prema kojoj su eukarioti (organizmi sa ćelijskom jezgrom) nastali kao rezultat niza asocijacija različitih stanica jedna na drugu. Moderni amandman na ovu teoriju kaže da formiranje eukariota, očigledno, nije bio opći trend koji je obuhvatio mnoge evolucijske grane (kako je Margulis pretpostavio), već jedinstven događaj koji je doveo do fuzije arhealnih i proteobakterijskih stanica. Kao rezultat toga, formirana je složena stanica s mitohondrijama, koja je postala prvi eukariot. Daljnji simbiogenetski događaji - na primjer, hvatanje algi koje su postale hloroplasti - dogodili su se mnogo puta, ali nisu povezani s pojavom eukariota kao takvih.
Prije više od pedeset godina, u martu 1967. godine, međunarodni časopis za teoretsku biologiju objavio je članak “O porijeklu ćelija koje se dijele mitozom” (L. Sagan, 1967. O porijeklu ćelija koje mitozuju). Autor članka se zvala Lynn Sagan, ali je ova izvanredna žena kasnije postala mnogo poznatija kao Lynn Margulis. Prezivala se Sagan jer je kratko bila udata za Carla Edwarda Sagana, astronoma i pisca.
Objavljivanje članka Lynn Margulis 1967. godine (nazvat ćemo je tako zbog pogodnosti) postalo je početak obnove bioloških koncepata, što su mnogi autori smatrali promjenom paradigme, odnosno, drugim riječima, pravom znanstvenom revolucijom. (I.M. Mirabdullaev, 1991. Endosimbiotička teorija - od fikcije do paradigme). Suština intrige ovdje je jednostavna. Još od vremena Charlesa Darwina, biolozi su uvjereni da je glavna metoda evolucije divergencija - divergencija grana. Lynn Margulis je bila prva koja je istinski objasnila naučnoj zajednici da je mehanizam nekih velikih evolucijskih događaja vjerovatno bio fundamentalno drugačiji. Margulisova interesovanja bila su usmerena na problem porekla eukariota - organizama čije ćelije imaju složenu unutrašnju strukturu sa jezgrom. Eukarioti uključuju životinje, biljke, gljive i mnoge jednoćelijske organizme - amebe, flagelate, cilijate i druge. Margulis je pokazao da se rana evolucija eukariota uopće nije svela na divergenciju - uključivala je spajanje evolucijskih grana, i to više puta. Činjenica je da najmanje dvije vrste eukariotskih organela - mitohondrije, zahvaljujući kojima možemo udisati kisik, i hloroplasti, koji vrše fotosintezu - ne potiču od istog pretka kao glavni dio eukariotske ćelije (Sl. 1) . I mitohondrije i hloroplasti su bivše bakterije, u početku potpuno nepovezane sa eukariotima (proteobakterije u slučaju mitohondrija i cijanobakterije u slučaju hloroplasta). Ove bakterije je apsorbirala stanica drevnog eukariota (ili pretka eukariota) i nastavile živjeti u njoj, zadržavši za sada svoj vlastiti genetski aparat.
Dakle, eukariotska ćelija je, kako Margulis kaže, multigenomski sistem. A nastao je kao rezultat simbioze, odnosno obostrano korisne kohabitacije različitih organizama (točnije, endosimbioze, čiji jedan od sudionika živi unutar drugog). Odgovarajuće evolucijske grane su se, naravno, spojile. Ovaj pogled na evoluciju naziva se teorijom simbiogeneze.
Sada je teorija simbiogeneze općenito prihvaćena. Ona je potvrđena striktno kao što se može potvrditi bilo koja teorija o evoluciji velikih razmjera. Ali naučni koncepti, za razliku od religijskih dogmi, nikada ne ostaju statični. Naravno, cjelokupna slika simbiogeneze sada nam ne izgleda sasvim isto (a na nekim mjestima ni najmanje isto) kako ju je zamišljala Lynn Margulis prije pola stoljeća.
Logic classic
Na pedesetu godišnjicu objavljivanja čuvenog članka o simbiogenezi Časopis za teorijsku biologiju pripremio posebno izdanje u potpunosti posvećeno kreativnom naslijeđu Lynn Margulis. Ovo izdanje uključuje opsežan članak poznatog britanskog biohemičara i popularizatora nauke Nicka Lanea, koji upoređuje trenutno stanje pitanja porijekla eukariota s klasičnim idejama na tu temu. Lane ne sumnja da je Margulis bio u pravu u glavnim izjavama (u vezi porijekla mitohondrija i hloroplasta); u naše vreme, čini se, niko od ozbiljnih naučnika u to ne sumnja, jer su podaci molekularne biologije po ovom pitanju nedvosmisleni. Ali đavo, kao što znamo, živi u detaljima. U ovom slučaju, udubljivanjem u detalje, tamo možemo pronaći puno novih i zanimljivih stvari, i što je najvažnije, pobrinuti se da tema o porijeklu eukariota nije daleko od iscrpljenosti.
Počnimo s činjenicom da su se neke Margulisove privatne pretpostavke pokazale netačnim. To je normalno: s obzirom na ogromnu brzinu razvoja biologije, jednostavno je nevjerovatno da je apsolutno sve točno naslućeno u članku objavljenom prije pola stoljeća. Nove činjenice koje tada autoru nisu mogle biti poznate sigurno će unijeti neke korekcije. Ovo se dogodilo i ovdje. Prije svega, Margulis je insistirao na simbiotičkom porijeklu ne samo mitohondrija i hloroplasta, već i eukariotskih flagela. Ona je vjerovala da su preci flagela bili dugačke, spiralno uvijene, pokretne bakterije vezane za eukariotsku ćeliju, slične modernim spirohetama (vidi sliku 1). Nažalost, ova hipoteza nije dobila nikakvu molekularnu biološku potvrdu, a sada je više niko ne podržava.
U nekim je trenucima Margulis mogla biti u pravu (to nije zabranjeno ni prirodnim zakonima ni unutrašnjom logikom njezine vlastite teorije), ali je ipak, iz razloga van njene kontrole, promašila. Na primjer, vjerovala je da, budući da su mitohondrije potomci bakterija, prije ili kasnije biolozi će naučiti da ih uzgajaju u hranljivom mediju izvan eukariotskih ćelija - pa, kao obični mikrobi. Da je to moguće, bio bi idealan dokaz teorije simbiogeneze. Nažalost, moderne mitohondrije su u osnovi nesposobne za samostalan opstanak, jer je većina njihovih gena, tokom evolucije, migrirala u ćelijsko jezgro i tamo bila integrirana u genom eukariotskog „domaćina“. Sada se proteinski proizvodi ovih gena sintetiziraju izvan mitohondrija, a zatim se transportuju u njega pomoću posebnih transportnih sistema koji pripadaju eukariotskoj ćeliji. Geni koji ostaju u samim mitohondrijama uvijek su malobrojni – nisu dovoljni za održavanje života. Godine 1967. to još niko nije znao.
Međutim, sve je to, uglavnom, posebno. Razmišljanje Lynn Margulis bilo je sintetičko: ona nije bila ograničena na objašnjenja pojedinačnih činjenica, već je nastojala da ih kombinuje u integralni sistem koji opisuje evoluciju živih organizama u kontekstu istorije Zemlje (Sl. 2). Savremena naučna saznanja omogućavaju testiranje snage ovog sistema ideja.

Stablo i mreža
Sve je počelo sa kiseonikom. U drevnoj Zemljinoj atmosferi nije postojao molekularni kiseonik (O2). Tada su cijanobakterije, koje su prve savladale fotosintezu kisika, počele otpuštati ovaj plin u atmosferu (za njih je to jednostavno bio nepotreban nusproizvod). U međuvremenu, čisti kiseonik je veoma toksična supstanca za svakoga ko nema posebna biohemijska sredstva zaštite od njega. Nije iznenađujuće da su emisije kiseonika iz cijanobakterija zatrovale Zemljinu atmosferu i dovele do masovnog izumiranja. Počeo je “holokaust kiseonika” (L. Margulis, D. Sagan, 1997. Mikrokosmos: četiri milijarde godina mikrobne evolucije).
Ovdje je već potreban amandman. Mnogi moderni istraživači vjeruju da je prijelaz iz biosfere bez kisika u biosferu kisika zapravo bio mnogo postepeniji i manje destruktivan nego što sugeriraju spekulacije o "holokaustu kisika" (vidi, na primjer: "Veliki događaj kisika" na prijelazu arheja i proterozoika nije bio veliki, nije događaj, “Elementi”, 02.03.2014.). Štaviše, moguće je da je pojava slobodnog kiseonika još verovatnija povećana raznolikost mikroorganizama, jer je oksidacija niza minerala atmosferskim kiseonikom obogatila hemijski sastav životne sredine i stvorila nove ekološke niše (M. Mentel, W. Martin, 2008. Energetski metabolizam među eukariotskim anaerobima u svetlu hemije proterozojskog okeana ). Općenito, ideja o pojavi kisika u atmosferi kao jednokratnoj grandioznoj katastrofi koja je cijelu povijest Zemlje podijelila na "prije" i "poslije" sada se čini zastarjelom.
Na ovaj ili onaj način, nema sumnje da su alfa-proteobakterije najviše koristile obogaćivanjem naše planete kiseonikom. Naučili su da direktno koriste kiseonik za proizvodnju energije - i to sa velikom efikasnošću. Ali jednoćelijski preci eukariota nisu imali takvu sposobnost. Bili su anaerobni, odnosno nisu mogli da udišu kiseonik. Ali oni su bili grabežljivci koji su naučili da apsorbuju manje ćelije putem fagocitoze. I to im je dalo izvrsnu priliku: da uhvate neke bakterije, ne probavljajući ih, već ih „porobljavajući“ i prisvajajući proizvode njihovog metabolizma. Nakon što su apsorbirali alfa-proteobakteriju, primitivni eukarioti su mogli udisati kisik - tako su nastale mitohondrije. A nakon što je apsorbirala cijanobakteriju, mogla je fotosintetizirati - tako su nastali hloroplasti. Margulis je vjerovao da su se takvi događaji dešavali mnogo puta, prateći opći trend koji se pojavio. Ovo je takozvana skripta serijska endosimbioza.
Dakle, Margulis se ispostavlja da je u određenoj fazi razvoja života endosimbioza postala gotovo univerzalni obrazac. Zatim bi u osnovi evolucijskog stabla eukariota trebala postojati bukvalno čitava mreža evolucijskih grana, koje se ukrštaju jedna s drugom zbog endosimbiotskih događaja i „rastu“ u približno istom smjeru - u onom koji je diktirala kombinacija tadašnji spoljašnji uslovi sa strukturnim karakteristikama ćelija (slika 3, A).
Mora se reći da je do kraja 20. stoljeća u evolucijskoj biologiji (a posebno u paleontologiji) ideja da većina velikih evolucijskih događaja ima prirodni i sistemski karakter već stekla određenu popularnost. Takav događaj istovremeno pokriva mnoge evolucijske grane, u kojima se, pod utjecajem zajedničkog nasljeđa, paralelno javljaju približno iste karakteristike (vidi, na primjer: A. G. Ponomarenko, 2004. Artropodizacija i njene ekološke posljedice). Primjeri takvih događaja nazivali su se sisavizacija (podrijetlo sisara), angiospermizacija (podrijetlo cvjetnica), artropodizacija (podrijetlo člankonožaca), tetrapodizacija (podrijetlo kopnenih kralježnjaka), ornitizacija (podrijetlo ptica) i još mnogo toga . Činilo se da se formiranje eukariota - eukariotizacija - savršeno uklapa u ovu seriju.
Na primjer, Kiril Eskov u svojoj divnoj knjizi „Istorija Zemlje i života na njoj“ (napisanoj 1990-ih) kaže sljedeće: „Najvjerovatnije su različite varijante eukariotičnosti, odnosno unutarćelijske kolonije, nastale mnogo puta (jer Na primjer, postoji razlog vjerovati da su crvene alge, koje se po mnogim ključnim karakteristikama oštro razlikuju od svih drugih biljaka, rezultat takve „nezavisne eukariotizacije“ cijanobakterija)“ (K. Yu. Eskov, 2000. Istorija Zemlje i život na tome).
Nažalost, u odnosu na eukariote (sada ne raspravljamo o drugim primjerima "-acija"), savremeni podaci bacaju sumnju na ovaj prelijepi scenario.
Problem mitohondrija
Počnimo s činjenicom da je hipoteza o crvenim algama o kojoj govori Eskov sada zastarjela. Molekularne studije pokazuju da evolucijska loza crvenih algi leži duboko unutar eukariotskog stabla (oni su prilično bliski srodnici zelenih biljaka), a njihova nezavisna eukariotizacija je krajnje malo vjerojatna.
Ali nešto drugo je mnogo ozbiljnije. Ako je simbiogeneza bio prirodan, dug, višestepeni proces, pa čak i tekao paralelno u različitim evolucijskim granama, onda bismo očekivali da ćemo vidjeti spektar prilično različitih prijelaznih stanja između eukariota i neeukariota. To je upravo ono što je Margulis mislio. Činjenicu da ova prelazna stanja nisu uočljiva, ona je (koliko se može suditi) smatrala čisto tehničkim problemom koji je povezan sa nedostatkom znanja i nesavršenim metodama. Da li je to istina sada kada znamo nemjerljivo više o živim ćelijama nego što smo znali prije pedeset godina?
Hajde da spekulišemo. Navodna serijska endosimbioza je trebalo da se odvija, prvo, postepeno, a drugo, malo drugačije u različitim evolucionim linijama (pošto u evoluciji nema točnih ponavljanja). Na osnovu toga, Margulis je predvidio da će prije ili kasnije biti otkriveni eukarioti koji imaju hloroplaste, ali nikada nisu imali mitohondrije; eukarioti koji su zadržali bakterijske flagele (koje se po strukturi oštro razlikuju od flagele eukariota); i konačno, prvenstveno anaerobni eukarioti, u čijim ćelijama nema tragova adaptacije na atmosferu kiseonika. Nijedno od ovih predviđanja nije potvrđeno. Nijedan od eukariota nema čak ni naznaku flagele bakterijskog tipa - njihova sredstva kretanja su potpuno drugačija. Nijedan od poznatih eukariota ne može se nazvati primarnim anaerobom - svi su, bez izuzetka, prošli kroz "fazu kisika" u nekom trenutku svoje evolucije. Konačno, svi eukarioti imaju ili aktivne mitohondrije, ili njihove ostatke koji su izgubili značajan dio svojih funkcija (hidrogenozomi, mitozomi), ili – u najgorem slučaju – mitohondrijske gene koji su uspjeli da se presele u jezgro.
Krajem 20. vijeka, postojala je popularna hipoteza da neki moderni jednoćelijski eukarioti nemaju i nikada nisu imali mitohondrije. Predloženo je da se takvi prvenstveno nemitohondrijski eukarioti rasporede u posebno carstvo Archezoa. Margulis je prihvatio ovu hipotezu prilično rano i bio joj vjeran do posljednjeg – čak i kada su je mnogi drugi naučnici već odbacili (L. Margulis et al., 2005. “Nesavršenosti i neobičnosti” u poreklu jezgra). Smatrala je vrlo vjerojatnim da prvenstveno nemitohondrijski eukarioti („arheoprotisti“) još uvijek žive u nekim nepristupačnim staništima bez kisika, gdje ih je vrlo teško otkriti. Nažalost, nikakvi „arheprotisti“ još nisu pronađeni, ali je pronađen bilo koji broj ostataka mitohondrija u onim jednoćelijskim organizmima koji su prethodno bili klasifikovani kao Archezoa. Trenutno je poznat samo jedan eukariot koji uopšte nema tragove mitohondrija - bičak Monocercomonoides, ali položaj ovog stvorenja na evolucijskom stablu ne ostavlja nikakvu sumnju da je ono nekada imalo mitohondrije (A. Karnkowska i sar., 2016. Eukariot bez mitohondrijske organele). Općenito, u ovom trenutku, bez izuzetka, svi slučajevi odsustva mitohondrija kod eukariota moraju se smatrati sekundarnim. To znači da u istoriji eukariota - barem njihovih modernih grupa - nije bilo drevne nemitohondrijske faze.
Margulis je vjerovala (sasvim razumno za svoje vrijeme) da je u određenom periodu u istoriji života eukariotizacija bila širok trend – „trend“, kako se sada kaže. Na temelju toga bilo bi moguće pretpostaviti da različiti eukarioti imaju različite pretke: na primjer, da su eukariotske alge evoluirale od cijanobakterija, životinje od predatorskih bakterija, a gljive od osmotrofnih bakterija koje apsorbiraju hranjive tvari preko površine stanice. Ova hipoteza nije u suprotnosti ni sa jednim fundamentalnim zakonima biologije. Ali, nažalost, to je upadljivo kontradiktorno činjenicama. Molekularna sistematika pokazuje da zajednički predak biljaka, životinja i gljiva nije bio prelazni oblik, već pravi eukariot, „potpuno razvijen“, kako to kaže Nick Lane. Možemo sa sigurnošću reći da je zajednički predak svih modernih eukariota već bila punopravna eukariotska stanica: imala je jezgro, endoplazmatski retikulum, Golgijev aparat, mikrotubule, mikrofilamente, mitohondrije i flagele. Općenito, kompletan skup eukariotskih karakteristika.
Imajte na umu da ovaj skup karakteristika ne uključuje hloroplaste. Nisu se pojavili kod svih eukariota i ne odmah. Osim toga, hloroplasti su sigurno stečeni više puta, i to na različite načine u različitim evolucijskim granama. Hloroplasti su slični primarni(kada eukariot napadne cijanobakteriju), i sekundarno(kada eukariot uhvati drugog eukariota sa cijanobakterijom unutra) pa čak tercijarni(kada jedan eukariot uhvati drugog eukariota, unutar kojeg živi treći eukariot, a unutar njega - cijanobakterija). Ovdje je evolucija, kako kažu, u punom jeku. Kod mitohondrija je situacija potpuno drugačija: na osnovu njihovog prisustva ne vidimo nikakvu posebnu raznolikost i nikakve prelazne faze (osim brojnih činjenica o sekundarnom gubitku, ali takve činjenice ne govore apsolutno ništa o porijeklu eukariota). Da je Margulisov scenario potpuno ispravan, onda bi situacija s mitohondrijama i flagelama bila otprilike ista kao s hloroplastima - ali to nije slučaj.
Ono u čemu je Margulis bio u pravu je da su eukarioti općenito prilično predisponirani da preuzmu endosimbionte. Ovdje možemo navesti razne primjere, uključujući stjecanje bakterijskih simbionta od strane nekih dubokomorskih crva, na kojima ti crvi zapravo žive (V.V. Malakhov, 1997. Vestimentiferanci su autotrofne životinje). Brza evolucija hloroplasta je najupečatljivija manifestacija ovog trenda. Samo "glumci" koji su ih stekli očigledno su već tada imali pun skup eukariotskih karakteristika, uključujući mitohondrije. Konfiguracija evolucijskog stabla eukariota, kakvog ga sada poznajemo, jednostavno ne dopušta druge verzije.
Na to, Lane dodaje da se osnovna struktura ćelija iznenađujuće malo razlikuje među različitim eukariotima, ovisno o njihovom načinu života (iako sam način života može jako varirati). Sve karakteristične komponente ćelije koje je čine eukariotskom općenito su raspoređene na isti način u biljkama, životinjama, gljivama, flagelatima i amebama... “Sada znamo da gotovo sve razlike između eukariota odražavaju sekundarne adaptacije.” , piše Lane u članku o kojem se raspravlja. Ujednačenost strukture eukariotske ćelije znači da prve faze njenog formiranja nisu ostavile praktički nikakve tragove u modernoj raznolikosti eukariota.
Jedinstveni događaj
Zaključci koje Lane danas donosi više se ne mogu nazvati novim ili neočekivanim. Savremeni podaci su najkompatibilniji sa pretpostavkom da je došlo do formiranja eukariotske ćelije pojedinačni događaj, završena (u vremenskoj skali koja nam je dostupna) vrlo brzo. Vjerovatno je da su preci eukariota prošli kroz svojevrsno usko grlo u ovoj fazi (u jednom ranijem članku, Lane je sugerirao da je to bila mala, nestabilna, kratkovječna populacija u kojoj su se dogodile sve glavne promjene; N. Lane, 2011. Energetika i genetika širom podjele prokariota i eukariota). Kao rezultat toga, nastao je prvi "potpuni" eukariot, čiji su se potomci raspršili u različite ekološke niše - ali se osnovna struktura ćelije više nije mijenjala. Dakle, nije bilo paralelne eukariotizacije. U svakom slučaju, moderna biologija ne nalazi dokaze koji to potvrđuju.
Podaci iz komparativne genomike sugeriraju da je granični događaj koji je odvojio eukariote od ostatka žive prirode bio sjedinjenje dviju stanica - arhealne (vjerojatno pripada jednoj od Lokiarchaeota) i bakterijske (vjerojatno pripada jednoj od Proteobacteria) . Dobiveni superorganizam postao je prvi eukariot (slika 3, B). Moderna „mainstream“ tačka gledišta identifikuje ovaj događaj sa akvizicijom mitohondrija (tzv. „rani mitohondrijski” scenario; videti, na primer: N. Yutin et al., 2009. Izvori fagocitoze i eukariogeneze). Zaista, mitohondrije su neosporni potomci proteobakterija i sigurno su kao simbionti prodrli u ćeliju arheje (ili primitivnog eukariota koji nije previše udaljen od arheje). Međutim, na pitanje kako su tačno stigli, Lane daje prilično neočekivan odgovor. Naime: "Ne znamo."
Sta je bilo? Prema klasičnoj teoriji, sve unutrašnje simbionte su eukariotske stanice stekle fagocitozom, odnosno hvatanjem pseudopodima uz izolaciju uhvaćenog objekta i njegovu kasniju probavu (u ovom slučaju neuspješna). Ovo je očigledno tačno za hloroplaste, ali vrlo sumnjivo za mitohondrije. Pretpostavka da se fagocitoza pojavila ranije od mitohondrija ne uklapa se dobro s bioinformatičkim podacima. Komparativna analiza proteinskih sekvenci pokazuje da su aktinski mikrofilamenti koji čine unutrašnji okvir bilo kojeg pseudopoda najvjerovatnije u početku bili nepokretni - proteini koji im također omogućavaju kontrakciju pojavili su se mnogo kasnije (E.V. Kunin, 2014. Logika slučaja). To znači da evolucija eukariota nije mogla započeti direktno fagocitozom – mitohondrije su stečene na neki drugi način.
No, mora se naglasiti da su sve ovo još uvijek samo nagađanja. Misterija porijekla mitohondrija, a da ne spominjemo porijeklo jezgra, još uvijek nije riješena.
Slučaj i nužnost
Dakle, da li je hipoteza o serijskoj endosimbiozi tačna? Da - u smislu da su se simbiotski događaji zaista dogodili mnogo puta u istoriji eukariota. To najbolje ilustruje duga, bogata i sada dobro proučena istorija hloroplasta (P. Keeling et al., 2013. Broj, brzina i uticaj endosimbioza plastida u evoluciji eukariota). Ne – u smislu da serijska endosimbioza nije bila preduslov za nastanak eukariota kao grupe. Endosimbiotski događaj koji je doveo do pojave eukariota bio je, koliko sada možemo suditi, jedinstven.
Dakle, scenario “paralelne eukariotizacije” nije potvrđen. To ne znači da se evolucijski događaji ovog tipa uopće ne događaju: neke od njih detaljno su opisali paleontolozi (na primjer, sisavljenje životinjskih gmazova, koji u nekoliko evolucijskih grana paralelno poprimaju karakteristike sisara) . Štaviše, lista ovakvih „paralelnih scenarija“ u poslednje vreme čak raste. “Elementi” su više puta pisali o hipotezi o samostalnom nastanku nervnog sistema u dvije potpuno različite grane višećelijskih životinja (vidi Rasprava o ulozi ctenofora u evoluciji se nastavlja, “Elementi”, 18.09.2015.). Ali pojava eukariota jedan je od najjedinstvenijih događaja u čitavoj istoriji života na Zemlji. Vjerovatno zbog toga ispada iz ove serije.
U savremenoj naučnoj literaturi postoji koncept kao hipoteza retkih zemalja(vidi hipotezu o rijetkim Zemlji). Zagovornici ove hipoteze priznaju da relativno jednostavan život (na bakterijskom nivou organizacije) može postojati na mnogim planetama i biti prilično čest u Univerzumu. Ali relativno složen život (eukariotski ili uporediv) nastaje samo pod najrjeđim spletom okolnosti; moguće je da postoji samo jedna planeta sa takvim životom u Galaksiji. Ako je hipoteza o rijetkim Zemljama tačna, onda je pojava eukariota najvjerovatnije prekretnica koja razdvaja “jednostavan” život (široko rasprostranjen) od “složenog” (malo vjerovatnog) života.
Autor čuvene knjige „Poreklo života“, Mihail Nikitin, nedavno je (i potpuno nezavisno) došao do sličnih zaključaka. „Još ne znamo koliko je prirodna bila pojava eukariota. Ako za druge faze razvoja života, kao što je prelazak iz svijeta RNK u svijet RNA-proteina, odvajanje prokariotskih ćelija iz predćelijskog „svijeta virusa” ili pojava fotosinteze, možemo sa sigurnošću reći da su prirodni i gotovo neizbježni, budući da se život već pojavio, onda bi pojava eukariota u prokariotskoj biosferi mogla biti vrlo malo vjerovatna. Moguće je da u našoj galaksiji postoje milijarde planeta sa životom na bakterijskom nivou, ali samo na Zemlji su se pojavili eukarioti, na osnovu kojih su se pojavile višećelijske životinje, a potom i inteligentna bića” (M. Nikitin, 2014. Nova hipoteza o porijeklu eukariotske ćelije). Možda nam je zbog toga tako teško razumjeti detalje porijekla eukariota: ovo je jedinstven (na planetarnoj skali) događaj, na koji je vrlo teško primijeniti princip uniformizma, koji zahtijeva „podrazumevano ” da polazi od jednoobraznosti faktora i procesa u svim vremenskim momentima. Ali upravo je to razlog zašto je misterija porijekla eukariota jedna od najfascinantnijih u cijeloj biologiji. Još uvijek ima mnogo neriješenih pitanja u ovoj oblasti; nisu sva spomenuta ovdje (kao u članku o kojem raspravlja Nick Lane).
Ministarstvo zdravlja Ukrajine
Državni medicinski univerzitet u Zaporožju
Zavod za mikrobiologiju, virusologiju i imunologiju
Sažetak na temu:
"Teorije o poreklu rikecija i mitohondrija"
pripremio student
3 kursa 30 grupa
Mikheeva Evgenia Sergeevna
Zaporozhye
1. Teorija simbiogeneze
2. Rickettsia
3. Otkriven najbliži srodnik mitohondrija
4. Nick Lane
5. Odlomci iz popularne naučne knjige Nika Lejna „Energija, seks, samoubistvo. Mitohondrije i smisao života"
6. Osobine metabolizma
Bibliografija
Teorija simbiogeneze
(simbiotska teorija, endosimbiotska teorija, teorija endosimbioze) objašnjava mehanizam nastanka nekih organela eukariotskih ćelija - mitohondrija, hidrogenosoma i plastida.
Teorija endosimbiotičkog porijekla hloroplasta prvi put je predložena 1883. Andreas Schimper, što je pokazalo njihovu samoreplikaciju unutar ćelije. Njegovom nastanku prethodio je zaključak A. S. Famintsina i O.V. Baranetsky o dvojnoj prirodi lišajeva simbiotskog kompleksa gljiva i algi (1867). K. S. Merezhkovsky je 1905. predložio sam naziv "simbiogeneza", po prvi put detaljno formulisao teoriju i čak na njenoj osnovi stvorio novi sistem organskog svijeta. Famintzin je 1907. godine, na osnovu Schimperovog rada, također došao do zaključka da su hloroplasti simbionti, poput algi u lišajevima.
Dvadesetih godina 20. stoljeća, teoriju je razvio B. M. Kozo-Polyansky, i sugerirano je da su mitohondrije također simbionti. Tada se dugo vremena simbiogeneza praktički nije spominjala u naučnoj literaturi. Proširena i konkretizirana teorija dobila je ponovno rođenje u djelima Lynn Margulis počevši od 1960-ih.
Simbiotsko porijeklo mitohondrija i plastida
Kao rezultat proučavanja slijeda baza u mitohondrijskoj DNK, dobijeni su vrlo uvjerljivi argumenti u prilog činjenici da su mitohondrije potomci aerobnih bakterija (prokariota) srodnih rikecija, koje su se nekada naselile u eukariotskoj stanici predaka i „učile“ da žive u njemu kao simbionti. Sada su mitohondriji prisutni u gotovo svim eukariotskim ćelijama; oni više nisu sposobni da se razmnožavaju izvan ćelije.
Postoje dokazi da originalni endosimbiotski preci mitohondrija nisu mogli ni uvoziti proteine ni izvoziti ATP. Vjerovatno su u početku primali piruvat iz ćelije domaćina, a korist za domaćina bila je neutralizacija kisika toksičnog za nukleocitoplazmu od strane aerobnih simbionta.
Simbiotska teorija porijekla organela postala je klasik moderne biologije. Na dnevnom redu su pitanja njegove konkretizacije, izgradnje filogeneze, simbioze, traženja porodičnih veza, općenito, velikog i važnog posla, bez kojeg je nemoguće „graditi most“ od teorije do prakse. Brojni podaci ukazuju da je taksonomsko porijeklo mitohondrija u redu Rickettsiales. Podsjetimo da rikecije, poput mitohondrija, ne mogu postojati izvan ćelije domaćina, ali za razliku od potonjih, često štete domaćinu, na primjer, uzrokujući tifus. Naš sunarodnik V.V. Emelyanov sa Instituta za epidemiologiju i mikrobiologiju im. N.F. Gamaleya i njegove kolege su još 2001. godine predložili hipotezu da moderne patogene rikecije i mitohondrije imaju zajedničkog pretka, slično endosimbionima sličnim rikecijama (RPE) koji koegzistiraju s paramecijom. Prema autorima rada (V.V. Emelyanov, M.Yu. Vysokikh), „poslednji zajednički predak ne samo da je izgubio suvišne gene..., već je i prenio neke vitalne gene u genom domaćina.” Koristeći imunoblotiranje proteina iz cijelih stanica i membranskih frakcija Rickettsia prowazekii (uzročnika tifusa), ustanovljeno je da je jedan od proteina kodiranih takvim geni protein vanjske membrane mitohondrija - porin, koji kodira nukleus, uvezen u mitohondrije, ali ga, funkcionalno, koristi i Rickettsia prowazekii. Ovo zapažanje je, ako ne dokaz, onda vrlo ozbiljna potvrda hipoteze.
Iako su rikecije po veličini usporedive s nekim virusima, jasno se razlikuju od njih. Ćelije rikecije sadrže i DNK i RNK (u omjeru 1:3,5); okruženi su staničnim zidom koji sadrži muramsku kiselinu i osjetljivi su na lizozim. Elektronske mikrofotografije ultratankih preseka pokazuju nuklearnu regiju i ćelijski zid.
Većina rikecija nikada nije uzgajana izvan žive ćelije, ali se mogu razmnožavati u inkubiranim jajima i u životinjskom tkivu; Iz žumančane vrećice kokošjeg jajeta možete dobiti 10 9 ćelija. U izoliranim ćelijama rikecije mogu se otkriti neki srednji metabolički enzimi. Tokom uzgoja, brzina metabolizma takvih stanica slabi, ali dodavanje ATP-a, organskih kiselina i aminokiselina ponovo stimulira njihovo disanje. Rikecije tako imaju svoj metabolizam; međutim, oni, vjerovatno zbog promjena u permeabilnosti površine ćelije, nisu u stanju regulisati unos i izlučivanje metabolita.
Povezane informacije.
Teorija simbiogeneze (simbiotička teorija, endosimbiotska teorija ili teorija endosimbioze) objašnjava mehanizam nastanka nekih organela eukariotske ćelije - mitohondrija, plastida i hidrogenozoma.
Suština koncepta je obostrano korisna koegzistencija organele sa ćelijom. Ovo sugerira endosimbiozu kao simbiozu koja je korisna za obje strane sa formiranjem eukariotskih ćelija (ćelija u kojima je prisutno jezgro). Zatim je korištenjem prijenosa genetskih informacija između bakterija proveden njihov razvoj i povećanje populacije. Prema ovoj verziji, sav daljnji razvoj života i životnih oblika je posljedica prethodnog pretka modernih vrsta.Teoriju o endosimbiotskom porijeklu hloroplasta prvi je predložio Andreas Schimper 1883. godine, koji je pokazao njihovu samoreplikaciju unutar ćelije.
Njegovom nastanku prethodio je zaključak A. S. Famintsyn i O. V. Baranetsky o dvojnoj prirodi lišajeva - simbiotski kompleks gljiva i algi (1867).
Jasne odredbe sistema sastavio je ruski botaničar i zoolog K. S. Merežkovski.
On je 1905. godine predložio sam naziv "simbiogeneza", po prvi put detaljno formulisao teoriju i na njenoj osnovi stvorio novi sistem organskog svijeta. Famintsyn je 1907. godine, na osnovu Schimperovog rada, također došao do zaključka da su hloroplasti simbionti, poput algi u lišajevima.
Dvadesetih godina 20. stoljeća, teoriju je razvio B. M. Kozo-Polyansky, i sugerirano je da su mitohondrije također simbionti. Tada se dugo vremena simbiogeneza praktički nije spominjala u naučnoj literaturi. Proširena i konkretizirana teorija dobila je ponovno rođenje u radovima Lynn Margulis, počevši od 1960-ih.
Kao rezultat proučavanja slijeda baza u mitohondrijskoj DNK, dobijeni su vrlo uvjerljivi argumenti u prilog činjenici da su mitohondrije potomci aerobnih bakterija (prokariota) srodnih rikecija, koje su se nekada naselile u eukariotskoj stanici predaka i „učile“ da žive u njemu kao simbionti. Sada su mitohondriji prisutni u gotovo svim eukariotskim ćelijama; oni više nisu sposobni da se razmnožavaju izvan ćelije.
Postoje dokazi da originalni endosimbiotski preci mitohondrija nisu mogli ni uvoziti proteine ni izvoziti ATP. Vjerovatno su u početku primali piruvat iz ćelije domaćina; korist za domaćina bila je neutralizacija kisika toksičnog za nukleocitoplazmu od strane aerobnih simbionta.
Plastidi, poput mitohondrija, imaju svoju prokariotsku DNK i ribozome. Očigledno, hloroplasti su nastali od fotosintetskih bakterija, koje su se svojevremeno naselile u heterotrofnim ćelijama protista i pretvorile ih u autotrofne alge.
Dokaz
Mitohondrije i plastidi:
Imaju dvije potpuno zatvorene membrane. U ovom slučaju, vanjski je sličan membrani vakuola, unutrašnji je sličan bakterijama,
Razmnožavaju se binarnom fisijom (i ponekad se dijele neovisno o diobi stanice), nikada se ne sintetiziraju de novo,
Genetski materijal - kružna DNK koja nije povezana s histonima (U smislu udjela GC, DNK mitohondrija i plastida je bliža DNK bakterija nego nuklearnoj DNK eukariota),
Imaju vlastiti aparat za sintezu proteina - ribozome i druge ribozome prokariotskog tipa - sa konstantom sedimentacije od 70S. Struktura 16s rRNA slična je strukturi bakterija.
Neki proteini ovih organela su po svojoj primarnoj strukturi slični sličnim proteinima bakterija i nisu slični odgovarajućim proteinima citoplazme.
Problemi
DNK mitohondrija i plastida, za razliku od DNK većine prokariota, sadrži introne.
Samo dio njihovih proteina je kodiran u vlastitoj DNK mitohondrija i hloroplasta, dok je ostatak kodiran u DNK ćelijskog jezgra. Tokom evolucije, dio genetskog materijala "protekao" je iz genoma mitohondrija i hloroplasta u nuklearni genom. Ovo objašnjava činjenicu da ni hloroplasti ni mitohondriji ne mogu više postojati (reproducirati) samostalno.
Pitanje porijekla nuklearno-citoplazmatske komponente (NCC), koja je zahvatila proto-mitohondrije, nije riješeno. Ni bakterije ni arheje nisu sposobne za fagocitozu, hraneći se isključivo osmotrofno. Molekularno biološke i biohemijske studije ukazuju na himernu arheo-bakterijsku prirodu JCC. Nije jasno kako je došlo do spajanja organizama iz dva domena.
Primjeri endosimbioza
Danas postoji niz organizama koji u svojim ćelijama sadrže druge ćelije kao endosimbionte. Oni, međutim, nisu primarni eukarioti koji su preživjeli do danas, u kojima se simbionti još nisu integrirali u jedinstvenu cjelinu i nisu izgubili svoju individualnost. Ipak, oni jasno i uvjerljivo pokazuju mogućnost simbiogeneze.
Mixotricha paradoxa je najzanimljiviji organizam sa ove tačke gledišta. Za kretanje koristi više od 250.000 bakterija Treponema spirochetes pričvršćenih na površinu njegove ćelije. Mitohondrije ovog organizma su izgubljene po drugi put, ali unutar njegove ćelije postoje sferne aerobne bakterije koje zamenjuju ove organele.
Amebe iz roda Pelomyxa također ne sadrže mitohondrije i stvaraju simbiozu s bakterijama.
Cilijati iz roda Paramecium stalno sadrže alge unutar svojih ćelija, posebno Paramecium bursaria formira endosimbiozu sa zelenim algama iz roda Chlorella.
Jednoćelijska flagelirana alga Cyanophora paradoxa sadrži cijanele, organele koje podsjećaju na tipične hloroplaste crvenih algi, ali se od njih razlikuju po prisutnosti tankog ćelijskog zida koji sadrži peptidoglikan. Veličina genoma cijanele je ista kao kod tipičnih hloroplasta i mnogo puta manja od veličine cijanobakterija.
Hipoteze za endosimbiotičko porijeklo drugih organela
Endosimbioza je najšire prihvaćena verzija porijekla mitohondrija i plastida. Ali pokušaji da se na sličan način objasni porijeklo drugih organela i ćelijskih struktura ne nalaze dovoljno dokaza i nailaze na opravdanu kritiku.
Peroksizomi
Christian de Duve je otkrio peroksizome 1965. Također je sugerirao da su peroksizomi bili prvi endosimbionti eukariotske ćelije, omogućavajući joj da preživi sa sve većom količinom slobodnog molekularnog kisika u Zemljinoj atmosferi. Peroksizomi, međutim, za razliku od mitohondrija i plastida, nemaju ni genetski materijal ni aparat za sintezu proteina. Pokazalo se da se ove organele formiraju de novo u ćeliji u ER i nema razloga da se smatraju endosimbiotima.