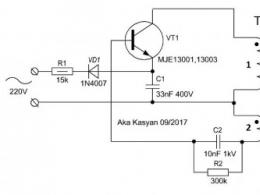
Классификация практическая и медицинское значение водоросли. Отдел водоросли
Учебное пособие написано в соответствии с требованиями ФГОС ВПО по направлению «Педагогическое образование» и дополняет знания студентов по теоретической части курса «Ботаника» (систематика растений и грибов). Материал пособия может быть использован студентами как для самостоятельной работы, так и для работы в аудитории под руководством преподавателя.
* * *
Приведённый ознакомительный фрагмент книги Ботаника. Систематика растений: учебное пособие (С. К. Пятунина, 2013) предоставлен нашим книжным партнёром - компанией ЛитРес .
Водоросли (Algae)
Многочисленная и разнообразная группа низших талломных растений, первичной средой обитания которых является вода. Водоросли объединяют несколько самостоятельных и, по всей вероятности, независимо эволюционировавших отделов. Представители отделов отличаются по набору пигментов, деталям тонкой структуры хроматофоров, по продуктам фотосинтеза, накапливающимся в клетке (запасным веществам), и по строению жгутикового аппарата. Низшие растения – одноклеточные, колониальные или ценобиальные и многоклеточные организмы. Ценобиями называют колонии, в которых число клеток определяется на ранних стадиях развития и не меняется до следующей стадии репродукции (воспроизведения). Рост ценобия происходит за счет увеличения размеров клеток, а не их числа. Различают следующие типы морфологической организации таллома:
1. Монадный – клетки, активно двигающиеся с помощь жгутиков.
2. Коккоидный – неподвижные клетки.
3. Ризоподиальный (амебоидный) – вегетативные клетки не покрыты оболочками и могут развивать цитоплазматические отростки – ризоподии.
4. Пальмеллоидный, или капсальный, тип организации представлен неподвижными клетками, погруженными в общую слизь.
5. Нитчатый – клетки, соединены в нити, простые или разветвленные.
6. Гетеротрихальный, или разнонитчатый, – усложненный вариант нитчатого строения, для которого характерны две системы нитей: стелющиеся по субстрату и отходящие от них вертикальные нити.
7. Пластинчатый – талломы в виде пластинок.
8. Сифональный – талломы, часто крупных размеров, формально представляют собой одну клетку обычно с большим числом ядер.
9. Сифонокладальная организация представлена многоядерными клетками, соединенными в нитчатые или иной формы многоклеточные талломы. На первых этапах образования таллома имеет сифональный тип строения.
Водоросли могут размножаться тремя способами: вегетативным, бесполым и половым. Вегетативное размножение заключается в отделении от целого растения части вегетативного таллома, дающей начало новому таллому. Бесполое размножение осуществляется при помощи специализированных клеток – спор, образующихся в спорангиях. Споры бывают подвижными (зооспоры) или неподвижными (апланоспоры). По форме они могут быть идентичны родительским талломам (автоспоры одноклеточных водорослей) или резко от них отличаться (одноклеточные споры многоклеточных водорослей).
Половое воспроизведение у водорослей чрезвычайно разнообразно. Наиболее простые формы полового процесса – слияние морфологически неразличимых вегетативных особей – хологамия и конъюгация. У значительной части водорослей происходит образование специализированных половых клеток – гамет. Различают следующее поведение гемет:
1. Изогамию - слияние одинаковых по форме и размерам гамет.
2. Гетерогамию – оба типа копулирующих гамет имеют жгутики, но женская крупнее и менее подвижна, чем мужская.
3. Оогамию - слияние неподвижной женской яйцеклетки и подвижной мужской клетки. Копулируют гаметы, возникшие на одной особи (гомоталлизм) или на разных особях (гетероталлизм). Гетероталлизм наблюдается при любой форме полового процесса. У изогамных форм гаметы при морфологическом тождестве оказываются физиологически различными и обозначаются условными знаками «+» и «-». Мужские гаметы, имеющие жгутики, называют сперматозоидами, не имеющие жгутиков, но способные передвигаться при помощи амебоидных движений называют спермациями. В результате полового процесса образуется диплоидная клетка – зигота.
Жизненный цикл, или цикл воспроизведения водорослей, включает вегетативный рост, бесполые спороношения, половой процесс, покоящиеся стадии. Соотношение диплоидной и гаплоидной фаз в жизненном цикле водорослей неодинаково. В одних случаях прорастание зиготы сопровождается редукционным делением (мейозом) зиготы (зиготическая редукция), при этом развивающиеся растения оказываются гаплоидными. У многих зеленых водорослей зигота – единственная диплоидная стадия в цикле развития, вся вегетативная фаза проходит у них у гаплоидном состоянии. Такой жизненный цикл называют моногаплобионтным. У части других водорослей, наоборот, вся вегетативная фаза диплоидна, гаплоидная фаза представлена лишь гаметами, перед образованием которых и происходит редукционное деление ядра (гаметическая редукция), жизненный цикл – монодиплобионтный. У третьих редукционное деление ядра предшествует образованию спор, развивающихся на диплоидных талломах (спорическая редукция). Они вырастают в гаплоидные растения размножающиеся половым путем (гаметофиты). После слияния гамет зигота развивается в диплоидное растение, несущее органы бесполого размножения (спорофиты ). Таким образом, у этих водорослей имеет место чередование поколений (генераций): диплоидного и гаплоидного. Жизненный цикл – гаплоидно-диплоидный. Оба поколения могут быть одинаковы морфологически (изоморфная смена поколений) или же резко отличаться по внешнему виду (гетероморфная смена поколений).
На практических занятиях изучают отделы: зеленые (Chlorophyta), диатомовые (Bacillariophyta или Diatomeae), бурые (Phaeophyta), красные (Rhodophyta) водоросли.
Отдел Зеленые водоросли (Chlorophyta)
Отдел Зеленые водоросли – самый большой по количеству видов (до 20 000 видов) и морфологически разнообразный отдел водорослей. Здесь имеются и микроскопические мелкие, одноклеточные формы (монадные и коккоидные) и достаточно сложно устроенные нитчатые, гетеротрихальные, сифональные, сифонокладальные и пластинчатые, достигающие нескольких десятков сантиметров. Обширна и область распространения зеленых водорослей (они встречаются по всему земному шару) и широка их экологическая амплитуда. Они обитают в пресных и морских водоемах, некоторые живут вне воды. Но при всем многообразии у зеленых водорослей есть ряд общих признаков:
1) пигментный состав: хлорофилл а и в, каротиноиды и ксантофиллы;
2) основной запасной продукт углеводной природы – крахмал, откладывается в хроматофоре вокруг пиреноида;
3) светочувствительный глазок – стигма, находится в строме хроматофора;
4) тилакоиды, несущие пигменты имеют тенденцию к образованию стопок;
5) жгутики изоморфные (одинаковые по строению) и изоконтые (равные по длине).
Класс Собственно зеленые, или Равножгутиковые, водоросли (Chlorophyceae, Isocantae)
Для представителей этого класса характерно бесполое размножение при помощи неподвижных апланоспор или подвижных зооспор с двумя – четырьмя, реже многими изоконтными и изоморфными жгутиками. Половые процессы – хологамия или копуляция гамет – изогамия, гетерогамия, оогамия. Зигота обычно проходит состояние покоя и прорастает при наступлении благоприятных условий, причем ее диплоидное ядро сразу делиться редукционно. В соответствии со ступенями морфологической дифференцировки таллома класс делится на порядки.
Порядок Вольвоксовые (Volvacales)
Порядок включает одноклеточные, колониальные и ценобиальные водоросли снабженные жгутиками, то есть монадной организации.
Род Хламидомонада (Chlamydomonas)
Хламидомонада – обширный род, объединяющий около 500 видов, широко распространен в природе. Его виды можно встретить в мелких, хорошо прогреваемых водоемах, лужах, канавах. При массовом развитии вызывает цветение воды, особенно в загрязненных органическими веществами водоемах. Таллом хламидомонады одноклеточный, монадной организации, то есть, находясь в активном состоянии, хламидомонады быстро передвигаются с помощью двух равных жгутиков, прикрепленных к переднему концу тела. Фаза активного движения сменяется состоянием покоя. Это так называемая пальмеллевидная стадия, когда клетки теряют жгутики, их оболочки сильно ослизняются и образуют агрегации клеток хламидомонад, погруженных в общую слизь. В таком виде клетки хламидомонад размножаются делением. Попадая в благоприятные условия существования, хламидомонады снова вырабатывают жгутики и переходят к активному движению.
Хламидомонада имеет целлюлозно-пектиновую клеточную оболочку, чашевидный хроматофор с одним или несколькими пиреноидами, расположенными в нижней части, и светочувствительным глазком (стигмой) – в верхней части. Ядро находится в углублении хроматофора, имеется пара пульсирующих вакуолей. Бесполое размножение зооспорами происходит в благоприятных условиях обитания. Каждая хламидомонада потенциально может размножаться как вегетативно, так и бесполым путем, а также участвовать в половом процессе. При бесполом размножении протопласт делится на 4 или 8 частей, образуются зооспоры. Половой процесс у большинства видов изогамный. Гаметы образуются так же, как и зооспоры, но в большем количестве (32 или 64). Зигота хорошо приспособлена к перенесению неблагоприятных условий. Ее прорастание сопровождается редукционным делением. Цикл развития хламидомонад – моногаплобионтный.
Род Вольвокс (Volvox)
Род Вольвокс – колониальные или ценобиальные водоросли. Небольшой род вольвокс обитает в чистых стоячих водоемах, прудах и небольших озерах. Это наиболее высокоорганизованный представитель порядка вольвоксовых. Он представляет собой крупный, достигающий 2–3 мм в диаметре шар, одетый тонким слоем слизи (инволюкрумом), под которым по периферии шара в один слой располагаются двужгутиковые клетки. Число их колеблется от 500 до 60 000. Внутренняя полость шара занята жидкой слизью. Клетки колонии по строению подобны клеткам хламидомонады. Но оболочка каждой клетки сильно ослизнена, поэтому протопласты соседних клеток удалены друг от друга и цитоплазматические отростки пронизывают толщу ослизненной оболочки. В местах соприкосновения формируются плазмодесмы.
В бесполом размножении участвуют 8-10 клеток, расположенных в задней, относительно направления движения, части сферы. Это гонидии. Среди других эти клетки выделяются более крупными размерами. При их делении сначала формируется плоская 16-клеточная пластинка (гоническая стадия), дальнейшее деление приводит к формированию незамкнутой сферы с маленьким незамкнутым отверстием, направленным к наружной поверхности родительской колонии. Формирующиеся клетки нового организма обращены своими жгутиками внутрь сферы. Нормальная ориентировка клеток (с передними концами, направленными кнаружи), достигается путем полного выворачивания наизнанку незамкнутой сферы, только после этого ее отверстие замыкается. Репродуктивные клетки дифференцируется очень рано, так что можно наблюдать внутри материнского организма не только дочерние, но внучатые колонии. Освобождаются молодые колонии после разрушения материнской.
Клетки, служащие для полового размножения, – оогонии и антеридии. Оогонии темно-зеленого цвета значительно крупнее других клеток и лишены жгутиков. В оогонии развивается одна крупная яйцеклетка. Антеридии формируют пакеты сперматозоидов. Половой процесс у вольвокса оогамный. Встречаются обоеполые и раздельнополые виды, а так же гомо-и гетероталличные клоны. Формируется покоящаяся зигота, которая прорастает молодой дочерней колонией после редукционного деления диплоидных ядер. Цикл развития моногаплобионтный.
Объекты: р. хламидомонада, р. вольвокс.
Ход работы
1. Рассмотрите хламидомонаду сначала на малом увеличении (м. увел.), затем более детально при большом увеличении (б. увел.) изучите неподвижные особи, пронаблюдайте движение хламидомонады.
2. Сделайте два рисунка:
а) внешний вид хламидомонады. Обозначьте оболочку, хроматофор;
б) схему строения клетки хламидомонады, используя таблицу. Обозначьте оболочку, цитоплазму, ядро, хроматофор, глазок (стигму), пиреноид, жгутики, пульсирующие вакуоли.
3. Рассмотрите при м. увел. и зарисуйте с препарата сферические ценобии вольвокса с дочерними колониями.
4. Сделайте схематичный рисунок, используя таблицу, отразив особенности строения ценобия. Обозначьте протопласты, цитоплазматические отростки, плазмодесмы, жгутики, оогонии, антеридии.
Порядок Хлорококковые (Chlorococcales)
Порядок включает одноклеточные и ценобиальные формы с коккоидной организацией клетки.
Род Хлорококк (Chlorococcum)
Род Хлорококк содержит 38 видов и встречается в разнообразных местообитаниях: в воде, как в планктоне, так и в бентосе; в почве, а также на коре деревьев, на старых деревянных постройках. Хлорококк входит в состав лишайников.
Это одноклеточная коккоидная водоросль, неподвижная в вегетативном состоянии. Клетки имеют чашевидный хроматофор с пиреноидом, но отсутствуют жгутики, глазки и пульсирующие вакуоли. Ядро располагается в выемке хроматофора. У старых особей можно наблюдать несколько ядер, клетки покрыты толстой целлюлозной оболочкой.
Размножается хлорококк бесполым путем с помощью двужгутиковых зооспор удлиненной формы. Протопласт материнской клетки делится и формируется от 8 до 32 зооспор, которые освобождаются после разрушения стенки материнской клетки. Период активного движения непродолжительный; поплавав некоторое время, зооспоры теряют жгутики, одеваются оболочкой, растут, достигая размеров, характерных для того или иного вида. Половой процесс изогамный. Цикл развития моногаплобионтный.
Род Гидродицион (Hydrodiction)
Гидродицион небольшой, но широко распространенный род. Встречается в заводях рек, прудов и других стоячих водоемах, обогащенных азотом. Это макроскопическая ценобиальная водоросль, сложенная из большого количества (до 20 000) клеток. Старые экземпляры достигают метровой длины, а клетки у них до полутора сантиметров.
Гидродикцион имеет вид замкнутой сети, состоящей из 5-6-угольных ячеек, образованных гигантскими клетками, соединенными своими концами. В клетках взрослых сеточек – одна гигантская вакуоль, цитоплазма располагается постенно и содержит сетчатый хроматофор с многочисленными пиреноидами и большое количество мелких ядер. Оболочка целлюлозная. Каждая клетка выполняет все функции организма (питание и размножение).
Размножение водяной сеточки бесполое и половое. К бесполому размножению приступают клетки, достигшие уже достаточно больших размеров (0,2 мм). В протопласте формируется такое количество зооспор, которое характерно для данного вида сеточки. Зооспоры не покидают материнскую клетку, а некоторое время перемещаются внутри клетки.
Затем сбрасывают жгутики и складываются в молодую сеточку, склеиваясь друг с другом теми местами, где проходят тяжи микротрубочек. Клетки молодой сеточки одноядерные, с пластинчатым хроматофором, несущим один пиреноид. Некоторое время молодая сеточка живет под оболочкой материнской клетки, но ее размер быстро увеличивается, клетки вытягиваются. В конце концов, оболочка материнской клетки разрушается, и сеточка приступает к самостоятельной жизни.
Половое размножение изогамное, гамет формируется больше, чем зооспор, и они значительно меньше. Зигота окрашена гематохромом в кирпично-красный цвет. После периода покоя зигота делится редукционно и прорастает четырьмя крупными зооспорами. Они малоподвижны, скоро теряют жгутики и снова покрываются толстой, но уже скульптурной оболочкой, превращаясь в так называемый полиэдр. Полиэдры, оболочки которых несут отростки, а содержимое богато жировыми включениями, по-видимому, имеют значение для распространения водорослей. Полиэдры могут благополучно переносить высыхание, и являются, таким образом, второй покоящейся стадией в цикле развития водяной сеточки.
Род Хлорелла (Chlorella)
Хлорелла – очень широко распространенная водоросль. В природе она встречается в планктоне и бентосе различных водоемов, на почве, участвует в образовании тела лишайников, а также живет в симбиозе с мелкими животными, образуя так называемые зоохлореллы. Это одна из культивируемых водорослей. Благодаря высокой скорости размножения, хлорелла дает большой выход биомассы.
Хлорелла – это одноклеточная коккоидная водоросль шаровидной формы. В протопласте находится колоколообразный, с большим углублением, хроматофор. Во впадине хроматофора можно обнаружить ядро. Размножается хлорелла только бесполым путем – автоспорами. Половой процесс неизвестен. Цикл развития бесполый, моногаплобионтный.
Объекты: р. хлорококк, р. водяная сеточка, р. хлорелла.
Ход работы
1. Рассмотрите при б. увел. и зарисуйте хлорококк. Обозначьте толстую оболочку (приспособление к аэрофильному образу жизни), чашевидный хроматофор.
2. Рассмотрите при м. увел. фрагмент ценобия водяной сеточки и зарисуйте, показав 5-6-угольные клетки, образующие ценобий. Обозначьте клетку ценобия, хроматофоры.
3. Рассмотрите при б. увел. и зарисуйте клетку ценобия с молодым ценобием внутри клетки. Обозначьте оболочку материнской клетки, молодой ценобий.
4. Зарисуйте хлореллу при б. увел. Обозначить толстую оболочку, хроматофор.
Порядок Улотриксовые (Ulothrichales)
Порядок объединяет водоросли, имеющие таллом в виде неразветвленной нити, сложенной из одноядерных клеток, реже таллом пластинчатый или трубчатый.
Род Улотрикс (Ulothrix)
Улотрикс – достаточно крупный род, встречается в пресных и чуть солоноватых водоемах, предпочитает чистую проточную воду. В прибрежной зоне рек и ручьев, особенно на территориях с подходящим климатом можно встретить Ulothrix zonata. Улотрикс бентосная, прикрепленная водоросль, образует скопления на прибрежных камнях.
Улотрикс – это многоклеточная нитчатая неветвистая водоросль. Все клетки, за исключением базальной, служащей для прикрепления, однотипны. Они покрыты тонкой, иногда ослизненной, целлюлозной оболочкой. Хроматофор постенный в виде замкнутого или незамкнутого пояска с большим количеством пиреноидов. Ядро в клетках одно. Число клеток в нитях неопределено, так как в верхних частях клетки постоянно делятся в одной плоскости. Размножается улотрикс вегетативно (фрагментацией нити), бесполым и половым путем. Бесполое размножение осуществляется зооспорами, снабженными четырьмя изоконтными и изоморфными жгутиками. Зооспоры, прорастая, формируют нити улотрикса. Зооспорангием потенциально может стать любая, кроме базальной клетка нити. Улотрикс образует двужгутиковые изогаметы. После слияния формируется планозигота (подвижная клетка с четырьмя жгутиками), которая затем теряет жгутики и дифференцируется в одноклеточный своеобразный спорофит – расширенное покрытое толстой оболочкой тело на тонкой ножке. В таком покоящемся состоянии зигота-спорофит находится некоторое время, и затем после редукционного деления в ней формируются зооспоры числом 4-16. Такой жизненный цикл характерен для Ulothrix zonata. У некоторых видов зигота прорастает в диплоидную нить. Таким образом, цикл развития у Ulothrix zonata – c гетероморфной сменой многоклеточного нитчатого гаплоидного спорогаметофита и одноклеточного диплоидного спорофита.
Род Ульва (Ulva)
Род Ульва известен под названием «Морской салат», широко распространен, но предпочитает мелкие воды. Ульва – морская водоросль, но хорошо переносит опреснение. Она обитает в устьях рек, мелководных лиманах, морских болотах; хорошо переносит, а отчасти и предпочитает воды со значительным органическим загрязнением. Местное население использует ее в пищу, но промыслового значения ульва не имеет.
Таллом ульвы – многоклеточный, пластинчатый, состоит из двух слоев клеток, края пластинки, гофрированные вследствие более интенсивных клеточных делений в краевых, по сравнению со срединными, зонах. В основании пластинка сужается в короткий черешок с подошвой, которой она и прикрепляется к твердому субстрату. Дифференциация клеток в талломе ульвы невелика. Специальных репродуктивных органов нет. Потенциально каждая клетка может стать спорангием у диплоидного поколения либо гаметангием у гаплоидного. Некоторые клетки имеют трубчатые выросты, спускающиеся вдоль центральной части таллома.
Бесполое размножение осуществляется четырехжгутиковыми зооспорами, образующимися на диплоидных растениях после редукционного деления. Прорастают зооспоры в однорядную неветвящуюся нить, но еще до начала деления у потерявшей жгутики и осевшей на грунт зооспоры обнаруживается поляризация. Верхний конец – более толстый, а нижний – тонкий и вытянутый, из него впоследствии формируются структуры прикрепления. Половой процесс изо– или гетерогамый, гаметы двужгутиковые. Цикл развития гаплодиплобионтный с изоморфной сменой поколений.
Объекты: р. улотрикс, р. ульва.
Ход работы
1. Рассмотрите при б. увел. и зарисуйте участок нити улотрикса, обратить внимание на строение хромотофора. Обозначьте клетки таллома: оболочку клетки, хроматофор, пиреноиды.
2. Зарисуйте внешний вид таллома ульвы, используя влажный препарат.
Порядок Хетофоровые (Chaetophorales)
Порядок включает многоклеточные нитчатые формы гетеротрихального типа с дифференцировкой таллома на горизонтальную, распростертую по субстрату, и вертикальную систему нитей.
Род Драпарнальдия (Draparnaldia)
Виды этого рода требовательны к чистоте и аэрации водоемов и предпочитают быстротекущие реки и ручьи. Они в массе произрастают на довольно значительных глубинах (10 м), образуя там целые заросли. Драпарнальдия бентосная – прикрепленная водоросль нитчатого гетеротрихального строения. У нее имеются длинные (неограниченного роста) слабоветвящиеся нити с поясковидным хроматофором с изрезанными краями. Хроматофор у таких веток небольшой относительно общего объема клетки, поэтому клетки магистральных нитей бледные. От этих нитей отходят мутовками пучки коротких сильноразветвленных веточек ограниченного роста, это ассимиляторы. Хроматофоры в них постенные, большие; клетки зеленого цвета. Каждая короткая нить заканчивается бесцветным длинным волоском. Органы размножения размещаются среди ассимиляторов. Бесполое размножение при помощи четырехжгутиковых зооспор. Половой процесс – изогамия или гетерогамия. Цикл развития моногаплобионтный.
Род Трентеполия (Trentepohlia)
Трентеполия – аэрофильная наземная водоросль, хорошо приспособленная к недостатку влаги. Она поселяется на коре деревьев, камнях, деревянных строениях. Особенно много видов этого рода обнаружено во влажных тропических и субтропических областях, где трентеполия нередко ведет эпифитный образ жизни. Трентеполию легко распознать среди других наземных водорослей за счет гематохрома – маслорастворимого каротиноида, придающего ей кирпичнокрасный или желтый цвет. Таллом трентеполии нитчатый гетеротрихальный. Стелющиеся по субстрату нити состоят из округлых или овальных клеток, покрытых толстой слоистой оболочкой. Они связаны друг с другом порами с плазмодесмами, но тем не менее стелющиеся нити легко распадаются на короткие фрагменты или отдельные клетки. Эти фрагменты и клетки в сухом состоянии распыляются и переносятся ветром. Таким образом, трентеполия имеет достаточно эффективный механизм вегетативного размножения, что особенно важно в условиях недостатка воды. В клетках находится нескольких дисковидных или ленточных хроматофоров, лишенных пиреноидов, и большое количество ядер, особенно у старых клеток. Кроме горизонтальной, имеется система вертикальных нитей, состоящая из более удлиненных клеток. Как горизонтальные, так и вертикальные нити обильно ветвятся за счет деления верхушечных клеток, на последних образуются слоистые колпачки.
Бесполое размножение происходит двух– или четырехжгутиковыми зооспорами, образующимися в специальных верхушечных клетках, – спорангиях, сидящих на трубчатых клетках – ножках. Спорангии отделяются и целиком переносятся воздушными течениями. Зооспоры образуются только в том случае, если спорангий окажется в воде. Тогда многоядерное содержимое очень быстро, за несколько минут, распадается на одноядерные участки, и вырабатываются зооспоры. Гаметангии также отличаются морфологически от вегетативных клеток, но располагаются преимущественно на стелющихся нитях. В отличие от спорангиев, шаровидные гаметангии не имеют ножек. Гаметангии также переносятся воздушными течениями. Попадая в воду, они прорастают двужгутиковыми изогаметами. Однако копуляция происходит редко, и гаметы развиваются партеногенетически (без оплодотворения). В случае формирования зигот они после периода покоя прорастают зооспорами. Цикл развития моногаплобионтный.
Объекты: р. драпарнальдия, р. трентеполия.
Ход работы
1. Рассмотрите при м. увел. и зарисуйте участок гетеротрихального нитчатого талома драпарнальдии, отметьте особенности строения «стволовых» нитей и ассимиляторов, покажите отличие строения хроматофоров в клетках осевой нити и в клетках боковых веточек. Обозначьте осевую нить, нити-ассимиляторы.
2. Рассмотрите при б. увел. и зарисуйте клетку осевой нити и клетку нити-ассимилятора. Обозначьте оболочку клетки, хроматофор.
3. Рассмотреть при б. увел. и зарисовать участок нити трентеполии. Обозначьте слоистую оболочку, запасные вещества (в виде капель масла, окрашенных гематохромом), дисковидные хроматофоры.
Порядок Кладофоровые (Cladophorales)
Порядок включает водоросли неклеточного строения, нитчатые ветвистые, поделенные поперечными перегородками на неравноценные сегменты, каждый из которых содержит много ядер. Поперечные перегородки возникают независимо от деления ядер.
Род Кладофора (Cladophora)
Кладофора – очень большой и широко распространенный род, в основном морских, и отчасти пресноводных водорослей. Описано около 150 видов. Встречается на мелководьях в полосе прибоя, на скалах, вдающихся в море, в лагунах, прудах, озерах. Молодые растения прикреплены к грунту или к различным подводным предметам, но позднее отрываются и плавают, образуя большие скопления или жесткую тину; а также крупные (10–15 см в диаметре) шаровидные скопления. Таллом сильноветвистый, сифонокладальной структуры. Сегменты, образуемые поперечными перегородками, не являются клетками в настоящем смысле. Формирование поперечных перегородок качественно отличается от цитокинеза и не связано с делением ядер. Сегменты неодинаковы по размеру и содержат разное количество ядер. Сетчатый хроматофор также формируется путем соприкосновения и слияния свободных вначале хлоропластов. Внешняя оболочка толстая, целлюлозная, никогда не ослизняется, поэтому тина кладофоры жесткая и не скользкая.
Размножение бесполое и половое. Зооспорангием или гаметангием может стать любой сегмент водоросли. Четырехжгутиковые зооспоры выходят из спорангия через пору и прорастают сначала в пузыревидное тело, лишенное перегородок (сифональная стадия), позже возникают поперечные перегородки и происходит ветвление нитевидных структур. Половой процесс изогамный. Зигота прорастает в диплоидное растение. У большинства видов кладофоры, обитающих в морях, изоморфная смена поколений, в таком случае зооспоры образуются после редукционного деления (мейозоо-споры), но у пресноводных видов отмечен монодиплобионтный цикл, когда редукционное деление предшествует образованию гамет.
Объекты: р. кладофора.
Ход работы
Рассмотрите при б. увел. и зарисуйте участок ветвящегося таллома кладофоры с зооспорангиями. Обозначьте клетки таллома, одноклеточные зооспорангии.
Класс Конъюгаты (Conjugatophyceae)
У представителей этого класса особый тип полового процесса – конъюгация, жгутиковых стадий нет, бесполого размножения спорами нет.
Порядок Зигнемовые (Zygnematales)
Порядок Зигнемовые – нитчатые неветвистые многоклеточные водоросли, оболочка их ослизняющаяся, и поэтому они скользкие на ощупь. Зигота прорастает одним проростком, три остальные ядра, образовавшихся в процессе редукционного деления, отмирают.
Род Спирогира (Spirogyra)
Род Спирогира – один из самых крупных и широко распространенных по всему земному шару: он найден даже в Антарктиде. Редкая канава, лужа, пруд или озеро лишены слизистой на ощупь, плавающей на поверхности тины. Таллом спирогиры – нитчатый, неветвистый, все клетки в нити равноценны и однотипны. В клетке имеется один или несколько постенных, спиральных, лентовидных хроматофоров, с большим количеством пиреноидов, расположенных вдоль продольной оси. Края хроматофора неровные. В клетке имеется одна или несколько вакуолей с клеточным соком. В первом случае цитоплазма занимает постенное положение. При наличии нескольких вакуолей, кроме постенного слоя цитоплазмы, имеются цитоплазматические тяжи и центральный цитоплазматический мешочек, в котором находится крупное, хорошо видимое без окраски, ядро. Внутренний слой клеточной оболочки целлюлозный, наружный – пектиновый, обеспечивает ослизнение и образование студенистого покрова, придающего нитям шелковистость. У спирогир, обитающих в водоемах с сильным течением, вырабатываются различного рода ризоиды, удерживающие водоросль на месте. Спирогира, как и другие коньюгаты, не имеет в цикле развития жгутиковой стадии и не образует спор. Она размножается либо фрагментацией нитей вегетативно, либо половым путем. При конъюгации формируются многочисленные зиготы, которые после периода покоя прорастают одной нитью. Из четырех гаплоидных ядер, образующихся в процессе редукционного деления, три мелких отмирают, и остается одно крупное жизнеспособное ядро. Все питательные вещества, накопленные в зиготе, идут на формирования одного проростка. Зиготы хорошо приспособлены к перенесению неблагоприятных условий. Они покрыты толстой трехслойной скульптурированной оболочкой. Структура оболочки зигот является важным таксономическим признаком.
Объекты: р. спирогира.
Ход работы
Рассмотрите при м. увел. и зарисуйте внешний вид многоклеточного таллома спирогиры. Обозначьте клетку таллома.
1. Рассмотрите при б. увел. и зарисуйте клетку спирогиры. Обозначьте оболочку клетки, хроматофор, пиреноиды,
ядро, тяжи цитоплазмы.
2. Пронаблюдайте и зарисуйте разные стадии конъюгационного процесса.
Обозначьте стадии: образование конъюгационных отростков, сжатие и перетекание протопласта, образование зиготы.
Порядок Десмидиевые (Desmidiales)
Порядок Десмидиевые – одноклеточные организмы или нитевидные колонии. Клетка состоит из двух равных половинок – полуклеток. При вегетативном размножении каждая полуклетка достраивает вторую половинку.
Род Клостериум (Closterium)
Клостериум – это пресноводная бентосная водоросль, требующая хорошего освещения, она обитает в небольших водоемах, прудах, тихих заводях рек и в обрастаниях подводных предметов. При массовом развитии формируют слизистые скопления. Клостериум любит чистую воду, но переносит органические загрязнения, а порой встречается и в сточных водах. Это одноклеточная форма, ее серповидное тело состоит из двух симметричных половин – полуклеток. Ядро расположено в центре, в цитоплазматическом мешочке. Внешней перетяжки, свойственной другим десмидиевым водорослям, клостериум не имеет, но внутренняя структура отвечает особенностям представителей этого порядка. Клостериум имеет два одинаковых осевых хроматофора, своеобразного строения. От центрального стержня радиально отходят несколько пластинок так, что на поперечном срезе хроматофор имеет вид многолучевой звезды. Крупные пиреноиды расположены вдоль стержня или разбросаны беспорядочно у основания. Основание хроматофора, обращенное к центру клетки, – широкое, по концам клетки хроматофор конически сужается. На полюсах клетки находятся две небольшие вакуоли с клеточным соком, в который погружены маленькие кристаллики гипса, находящиеся в постоянном движении (броуновское движение), и особые слизистые тельца. Трехслойная оболочка клостериума пронизана многочисленными коническими порами. Особенно крупные поры располагаются на концах клеток. Эти поры выделяют слизь, благодаря которой водоросль медленно передвигается. В то время как один конец тела прикрепляется к субстрату, другой совершает колебательные движения. Затем водоросль прикрепляется к субстрату другим концом. Так кувыркаясь, клостериум передвигается по направлению к источнику света.
Размножение клостериума вегетативное. Клетки делятся поперечно. Каждая дочерняя клетка получает половину материнской с одним хроматофором. Вторая половинка, то есть полуклетка, достраивается заново. Сначала молодая полуклетка не имеет хлоропласта, и лишь затем хроматофор старой полуклетки делится, и одна его половина переходит в новую полуклетку. Таким образом, каждая особь клостериума состоит из двух разновозрастных половинок: одной более старой и другой более молодой. Половой процесс – конъюгация двух особей, погруженных в общую слизь. Зигота покрыта толстой слоистой оболочкой и хорошо приспособлена к перенесению неблагоприятных условий. Всю зиму зиготы находятся в состоянии покоя, и долгое время ядра остаются не слившимися. Из зиготы формируется два молодых клостериума, получающих по два гаплоидных мейотических ядра. Одно, маленькое, вскоре дегенерирует. Второе – становится ядром новой особи. Цикл развития моногаплобионтный.
Объекты: р. клостериум.
Ход работы
1. Рассмотрите клостериум при м. увел. и пронаблюдайте его движение.
2. Рассмотрите клетку клостериума при б. увел. и зарисуйте ее. Обозначьте оболочку клетки, ядро, хроматофоры, пиреноиды, терминальные вакуоли с кристаллами гипса.
Вопросы и задания
1. Перечислите основные типы морфологической организации у зеленых водорослей. Охарактеризуйте каждый из типов. Приведите примеры.
2. Какие признаки характерны для представителей отдела зеленых водорослей?
3. На какие классы подразделяют отдел зеленые водоросли? В чем отличие представителей каждого класса?
4. Что такое колонии и ценобии у зеленых водорослей? Приведите примеры колониальных и ценобиальных зеленых водорослей. В чем различия колониальных и многоклеточных организмов?
5. Как происходит вегетативное размножение у зеленых водорослей? Приведите примеры.
6. Как происходит бесполое размножение у зеленых водорослей? Приведите примеры.
7. Какие половые процессы характерны для зеленых водорослей? Приведите примеры.
8. Зарисуйте схематично жизненный цикл хламидомонды, улотрикса, ульвы, спирогиры. Подпишите на схеме какое поколение является спорофитом, а какое гаметофитом, набор хромосом (гаплоидный или диплоидный) для каждого поколения и клеток служащих для размножения, тип редукционного деления и полового процесса. Укажите характерный для данных водорослей тип жизненного цикла.
9. Заполните таблицу:
Сравнительная характеристика классов и порядков
10. Охарактеризуйте роль зеленых водорослей в жизни водоемов. Приведите примеры зеленых водорослей, ведущих наземное существование.
Отдел Диатомовые водоросли (Bacillariophyta, Diatomeae)
Обширный отдел одноклеточных и колониальных организмов, объединяющих более 10 000 видов. Клетки диатомовых покрыты кремнеземовым панцирем, состоящим из двух половинок, надевающихся друг на друга, как крышка на коробку. Большая половинка – эпитека и меньшая – гипотека. Каждая половинка состоит из створки (донышка) и спаянного с ней пояскового кольца (пояска). Причем поясок эпитеки накладывается на поясок гипотеки. Под панцирем находится пектиновая оболочка. Диатомовые относятся к группе буроокрашенных водорослей, для которых характерно наличие хлорофиллов а и с, маскирующихся желтым пигментом фукоксантином. Запасной продукт углеводной природы – хризоламинарин. Монадные клетки – сперматозоиды с одним перистым жгутиком. Размножаются диатомовые вегетативно, путем продольного деления клеток с достраиванием одной створки – гипотеки. Половые процессы – конъюгация и оогамия. После полового процесса формируется зигота, обладающая способностью к росту (ауксоспора). Диатомовые водоросли живут в диплоидном состоянии и только их гаметы гаплоидны.
Класс Центрические (Centrophyceae)
Класс Центрические объединяет водоросли с радиально симметричным панцирем и отсутствием шовно-узелкового аппарата. Все центрические водоросли неподвижны. Половой процесс – оогамия.
Род Мелозира (Melosira)
Мелозира – это нитчатая колониальная водоросль, состоящая из однотипных цилиндрических клеток, соединенных между собой соприкосновением мелких шипиков, расположенных на округлых поверхностях створок. Панцирь мелозиры имеет широкие пояски, поэтому чаще всего водоросль рассматривается со своей боковой поверхности. В клетках имеются несколько лопастных хроматофоров, расположенных постенно, центр клетки занят большой вакуолью с клеточным соком.
Мелозира не имеет шовно-узловой структуры и поэтому неподвижна.
Размножается делением и половым путем. Половой процесс – оогамия. В одних клетках после редукционного деления образуется одна яйцеклетка, в других – четыре сперматозоида с одним жгутиком. Зигота покрыта тонкой, хорошо растяжимой пектиновой оболочкой. Интенсивно растущие зиготы называются ауксоспорами. Так как в половом процессе участвуют измельченные после многократных делений клетки, то ауксоспора восстанавливает их исходный объем. Достигнув определенных размеров, ауксоспора вырабатывает собственный панцирь. Цикл развития мелозиры монодиплобионтный.
Объекты: р. мелозира.
Ход работы
1. Рассмотрите участок колонии мелозиры при б. увел. и зарисуйте водоросль со стороны пояска и со стороны створки. Обозначьте клетку колонии, эпитеку, гипотеку.
2. Найдите и зарисуйте ауксоспору.
Класс Пеннатные (Pennatophyceae)
Класс Пеннатные включает водоросли с двусторонней симметрией панциря, имеющие шовно-узелковый аппарат, обладают способностью передвигаться. Половой процесс – конъюгация.
Род Пиннулярия (Pinnularia)
Род Пиннулярия включает более 150 видов. Обитает в пресных, бедных известью, водоемах. Ведет бентосный образ жизни на дне или в обрастаниях подводных предметов. Пиннулярия, как и другие диатомовые водоросли, имеет большое значение как кормовая база мелких животных и является начальным звеном пищевых цепей в водных экосистемах. Это одноклеточная водоросль, имеющая шовно-узелковую структуру и вследствие этого – подвижная. Среди других одноклеточных диатомовых водорослей пиннулярия имеет крупные размеры, и поэтому удобна для изучения. С пояска панцирь имеет прямоугольные очертания, а створки – от линейных до эллиптических. Концы створок большей частью закругленные, но могут быть оттянутые и головчатые. В центре и на концах створки хорошо заметны узелки и два s-образных щелевидных отверстия (шов), идущих от периферических узелков к центральному. По краям створки хорошо заметен, особенно на пустых панцирях, четкий рисунок из параллельных перегородок – септ, не достигающих шва. Клетки пиннулярии одноядерные с двумя пластинчатыми хроматофорами, с загнутыми краями. Широкой плоской стороной хроматофор обращен в сторону пояска, а краями выходит на сторону створки. Живые активные клетки пиннулярии окрашены в желтовато-бурый цвет, так как фукоксантин маскирует зеленые пигменты, но у отмирающих клеток фукоксантин вымывается, и хроматофор становится зеленым. В клетках имеется две вакуоли, разделенные центральным цитоплазматическим мостиком. В нем заключено ядро. Пиннулярия запасает волютин, видимый в световой микроскоп как тускло блестящие шаровидные тельца, и капли масла. Под панцирем клетка одета ослизняющейся пектиновой оболочкой. Пиннулярия, обладающая шовно-узловым аппаратом, активно передвигается, ползая по субстрату.
Размножается пиннулярия делением, проходящим параллельно створкам. Каждая дочерняя створка получает одну материнскую створку, вторая же, достраивающаяся створка, всегда – гипотека. Вследствие такой особенности при каждом делении одна дочерняя клетка всегда несколько мельче материнской, и в популяции можно встретить особи разного размера. Половой процесс у пиннулярии не обнаружен, ауксоспор не образуется. Можно предположить, что крупные клетки делятся чаще, чем мелкие, а самые маленькие совсем не делятся. Жизненный цикл – монодиплобионтный.
Объект: р. пиннулярия.
Ход работы
1. Рассмотрите пиннулярию при б. увел. Зарисуйте клетку со стороны створки. Обозначьте оболочку клетки, шов, узелки, септы, хроматофор.
2. Зарисуйте клетку со стороны пояска. Обозначьте эпитеку, гипотеку.
Вопросы и задания
1. Какие уровни и типы морфологической организации характерны для представителей диатомовых водорослей?
2. Каковы особенности строения клеток диатомовых водорослей?
3. Какое строение имеет панцирь у диатомовых водорослей?
4. Каковы принципы классификации диатомовых водорослей?
5. Какие способы размножения характерны для диатомовых водорослей?
6. Как осуществляются половые процессы у диатомовых водорослей? Что такое ауксоспора?
7. Где обитают диатомовые водоросли? Какие черты приспособления к планктонному и бентосному образу жизни есть у диатомовых водорослей? Приведите примеры планктонных и бентосных диатомовых водорослей.
8. Зарисуйте схематично жизненный цикл пиннулярии и мелозиры. Подпишите на схеме, какое поколение является спорофитом, а какое гаметофитом, набор хромосом (гаплоидный или диплоидный) для каждого поколения и клеток служащих для размножения, тип редукционного деления и полового процесса. Укажите характерный для данных водорослей тип жизненного цикла.
Отдел Бурые водоросли (Phaeophyta)
Основными пигментами бурых водорослей являются хлорофиллы а и с, каротиноиды и ксантофиллы, в том числе и фукоксантин, маскирующий зеленые пигменты и придающий водорослям характерную бурую окраску. Запасные продукты – ламинарин (вещество углеводной природы); маннит – сахароспирт и в небольшом количестве жиры. Феопласты пластинчатые или чаще многочисленные дисковидные (зернистый хроматофор), расположенные в перинуклеарном пространстве под наружной мембраной ядра, которая охватывает каждый феопласт, формирует феопластную эндоплазматическую сеть. Пиреноиды выступают над поверхностью в виде почки. В клетках имеется одно ядро, крупные вакуоли с клеточным соком и мелкие вакуоли, содержащие дубильные вещества и называемые физодами.
Оболочка состоит из двух слоев: наружного ослизняющегося, содержащего альгиновую кислоту, и внутреннего, образованного особой разновидностью целлюлозы – альгулезой. Монадные клетки с двумя гетероконтными и гетероморфными жгутиками располагаются с боковой стороны клетки. Передний – длинный перистый, покрытый мастигонемами, задний – короткий гладкий. Диапазон морфологических структур большой: от нитчатых гетеротрихальных – до дифференцированных пластинчатых тканевых форм. У всех бурых водорослей, за исключением представителей порядка фукусовые, у которых отсутствует бесполое размножение и которые являются диплобионтами, наблюдается смена генераций: у одних – изоморфная, у других – гетероморфная. Эти разные типы жизненного цикла положены в основу современного деления отдела бурых водорослей на три класса.
За немногими исключениями бурые водоросли – морские, особенно богато представленные в холодных водах северного и южного полушария.
Класс Изогенератные (Isogeneratophyceae)
Класс Изогенератные включает водоросли с изоморфным чередованием поколений или с гетероморфным, но с доминированием гаметофита.
Порядок Эктокарповые (Ectocarpales) включает гетеротрихальные формы.
Род Эктокарпус (Ectocarpus)
Эктокарпус – широко распространенная морская, бентосная водоросль, встречающаяся во всех широтах в прибрежной полосе морей и океанов. Водоросль поселяется на камнях и других подводных предметах, в том числе и на растениях. Будучи толерантными к различной солености воды, эктокарпусы участвуют в обрастаниях судов. Широка и температурная амплитуда местообитаний эктокарпуса. Он растет как в холодных, так и в теплых морях, активно функционирует как летом, так и зимой. Эктокарпус – макроскопическая (до 60 см) водоросль, нитчатая гетеротрихальная, имеющая вид ветвистых кустиков.
В основании расположены горизонтальные стелющиеся ризоиды, прикрепляющие водоросль к субстрату. Вертикальные веточки в основании таллома, покрытые корой из ризоидальных нитей, к верхушке становятся тоньше и заканчиваются длинными бесцветными клетками. Нарастание идет за счет деления клеток, расположенных в разных частях таллома. В клетках эктокарпуса имеются небольшие вакуоли с клеточным соком, одно ядро в постенном слое цитоплазмы и несколько лентовидных хроматофоров с пиреноидами. В стареющих клетках хроматофор становится дисковидным.
Размножение эктокарпуса бесполое и половое. Мейозоо-споры, почковидной формы с двумя неравными жгутиками, прикрепляющимися к боковой поверхности, образуются в одноклеточных спорангиях, сидящих на одноклеточной ножке. Такие спорангии образуются на диплоидных растениях спорофитах. На гаплоидных растениях закладываются многогнездные репродуктивные органы. Каждое гнездо продуцирует одну репродуктивную клетку монадной организации. Чаще всего эти клетки ведут себя как гаметы и, сливаясь, дают зиготу. Они однотипны морфологически, но их поведение различается: физиологически женские гаметы менее подвижны и быстро оседают на дно; физиологически мужские – более активны. Из зиготы вырастает диплоидный спорофит. Описанный выше цикл соответствует дипло-гаплобионтному типу с изоморфным чередованием поколений, но у эктокарпуса имеются отклонения от этого цикла, например, гаметы могут прорастать партеногенетически без оплодотворения, давая новые гаплоидные особи. Таким образом, видимо, достигается возможно полная адаптация эктокарпуса к условиям существования и объясняется его широкая экологическая амплитуда.
Объекты: р. эктокарпус (гербарные экземпляры, микропрепараты).
Ход работы
1. Рассмотрите гербарные образцы эктокарпуса. Зарисуйте внешний вид гетеротрихального таллома эктокарпуса.
2. Рассмотрите при б. увел. и зарисуйте с препарата участки таллома эктокарпуса с многокамерными гаметангиями и однокамерными спорангиями. Обозначите клетки таллома, гаметангий, спорангий.
Класс Гетерогенератные (Heterogeneratophyceae)
Класс Гетерогенератные характеризуется гетероморфной сменой поколений с доминированием спорофита и микроскопическими мелкими гаметофитами.
Включает водоросли со сложно устроенным талломом, имеющим тканевую структуру. Нарастание осуществляется за счет деления клеток, расположенных в месте перехода листовидной части таллома в черешок (интеркалярный рост).
Род Ламинария (Laminaria)
Известны и широко распространены в северных морях два вида ламинарии: Л. сахарная (L.saccharina (L.) Lamour) и Л. пальчатая (L.digitata (Hudz.Lam)), которые в верхней сублиторали образуют целые плантации и являются промысловыми водорослями. Ламинария носит название «морская капуста» и представляет собой ценный пищевой продукт, используемый в кормлении животных, а также изготовления различных продуктов питания и лекарственных препаратов для человека.
Ламинария – крупная, сложно-дифференцированная бентосная прикрепленная водоросль, истинно тканевого строения. Ее меристематические клетки способны делиться в трех взаимно перпендикулярных направлениях и образовывать объемные структуры, в которых все клетки связаны друг с другом плазмодесмами. Таллом ламинарии дифференцирован на три четко различимые части: мощные когтевидные ризоиды, которыми она прикрепляется к субстрату, радиально симметричную часть так называемый черешок и уплощенную пластинку. Первые две структуры многолетние, а пластинка отмирает и снова нарастает благодаря интеркалярной меристеме, находящейся в верхней части черешка. Внутренняя организация тела ламинарии тоже достаточно сложна. Черешок с поверхности имеет плотно сомкнутые клетки, содержащие зерна феопластов. В более глубоких слоях клетки бесцветные и удлиненные в продольном направлении. В центре клетки расположены рыхло и образуют сердцевину. Черешок постепенно нарастает в толщину, и на поперечном срезе хорошо видны слои, напоминающие годичные кольца. В анатомическом строении пластинки тоже различают мелкоклеточную кору. В центре проходит «жилка», клетки которой напоминают по своему строению ситовидные клетки высших растений.
Размножаются ламинарии половым и бесполым путем. Спорангии располагаются на поверхности пластинки, образуя целые спороносные участки или поля, на которых палисадным слоем расположены мешковидные спорангии и удлиненные стерильные отростки-парафизы. В спорангиях после редукционного деления образуются двужгутиковые почковидные мейозооспоры, прорастающие в гаплоидные организмы. Женские и мужские гаметофиты отличаются от спорофита. Это микроскопические нитевидные, слаборазвитые растения (заростки). Они недолговечны, их основная функция – продукция гамет. Половой процесс – оогамия. Зигота без периода покоя прорастает и развивается в диплоидный спорофит. Таким образом, почти вся активная жизнедеятельность ламинарии происходит в диплоидном состоянии. Цикл развития – диплогаплобионтный с гетеротрофным чередованием поколений, с доминированием спорофита.
Конец ознакомительного фрагмента.
Зелёные водоросли - одноклеточные, колониальные и многоклеточные формы, разнообразного строения, зелёного цвета. Продукт ассимиляции - крахмал, мука, масло. Имеются как подвижные формы со жгутиками на переднем конце клеток, так и неподвижные, прикреплённые или пассивно плавающие. Размножение вегетативное, бесполое и половое. У ряда форм имеется чередование бесполого и полового размножения. Зооспоры и гаметы с 2 или 4 жгутиками, расположенными на переднем конце. Пресноводные и морские водоросли.
К отделу пирофитовых относятся весьма своеобразные, одноклеточные водоросли. Они представляют собой неоднородную группу. У громадного большинства представителей этого отдела в строении клеток отчётливо выражены спинная, брюшная и боковые стороны. Особо следует сказать о хлоропластах. По разнообразию их окраски пирофитовым принадлежит первое место среди водорослей. Обычно хлоропласты окрашены в оливковый, бурый или коричневый цвет. Громадное большинство пирофитовых водорослей характеризуется жгутиками. Продуктом ассимиляции является крахмал или масло, изредка лейкозин и валютин. Размножение в основном вегетативное. Реже наблюдается бесполое размножение. Половой процесс достоверно неизвестен. Пирофитовые водоросли широко распространены в водоёмах и обитают в пресных и солоноватых водах, как и в морях.
К отделу золотистых относятся водоросли, преимущественно микроскопические, хлоропласты которых окрашены в золотисто - жёлтый цвет. В зависимости от пигментов окраска водорослей может приобрести различные оттенки: от чисто- золотисто - жёлтой до зеленовато - жёлтой и золотисто - бурой. В процесс фотосинтеза в клетках золотистых водорослей вместо крахмала вырабатывается особый углевод - лейкоцин. Живут преимущественно в чистых пресных водах. Небольшое число их обитают в морях и солёных озёрах. Золотистые водоросли бывают одноклеточными, колониальными и многоклеточными. Многие виды снабжены жгутиками. Размножаются золотистые водоросли простым делением клетки. Наблюдается также бесполое размножение.
Диатомовые водоросли - это совершенно особая группа одноклеточных организмов, одиночно живущих или объединенных в колонии различного типа: цепочки, нити, ленты, звёздочки. Окраска хлоропластов у диатомовыз водорослей имеет различные оттенки жёлто - бурого цвета в зависимости от набора пигментов. В процессе фотосинтеза у диатомовых водорослей образуется масло в виде капелек различной величины. Чаще всего диатомовые размножаются вегетативным делением клетки на две половины. Большинство диатомовых водорослей передвигаются по субстрату толчками вперёд, назад и слегка в сторону. Диатомовые водоросли живут повсюду. Водная среда - основное и первичное их место обитания.
К отделу жёлто - зелёных водорослей относятся водоросли, хлоропласты которых окрашены в светло- или тёмно - жёлтый цвет, очень редко зелёный и лишь иногда голубой. Окраска эта определяется наличием в хлоропластах основного элемента - хлорофилла. Кроме того, у них в клетках отсутствует крахмал, а в качестве основного продукта ассимиляции накапливаются капли масла и лишь у некоторых глыбки лейкоцина и валюцина. Встречаются они главным образом в чистых пресноводных водоёмах, реже в солоноватых водах и морях.
Отличительной особенностью жёлто - зелёных водорослей является наличие жгутиков. Именно этот признак в своё время послужил основанием для того, чтобы назвать эту группу водорослей разножгутиковыми. Помимо различий в длине, жгутики здесь различаются и морфологически: главный жгут состоит из оси и перисто расположенных на ней мерцательных волосков, боковой жгут бичевидный. Размножаются жёлто - зелёные водоросли простым делением клетки или распадом колоний и многоклеточных талломов на отдельные части. Наблюдается также бесполое размножение. Половой процесс известен лишь у немногих видов.
Эвиленовые водоросли - обычные обитатели небольших пресных стоячих водоёмов. Форма тела эвиленовых водорослей хорошо приспособлена к движению в воде. Движение эвиленовых водорослей совершается с помощью жгута. Процесс размножения у эвиленовых водорослей наблюдается обычно вечером или рано утром. Заключается он в делении особи надвое.
Хоровые водоросли - представляют собой совершенно своеобразные крупные растения, резко отличающиеся от всех остальных водорослей. Они широко распространены в пресноводных прудах и озёрах, особенно с жёсткой известковой водой. Набор ассимиляционных пигментов сходен с таковым у зелёных водорослей. При размножении этих клеток их ядра делятся метотически. Количество выявляющихся при этом хромосом у разных видов различное, от 6 до 70.
Сине - зелёные водоросли - старейшая группа организмов. Сине - зелёные водоросли встречаются во всевозможных и почти невероятных для существования местах обитания, на всех континентах и во всевозможных водоёмах Земли. Окраска их варьируется от чисто-сине-зелёной до фиолетовой или красноватой, иногда до пурпурной или коричневато-красной. Самым обычным типом размножения у сине-зелёных водорослей является деление клеток надвое. Сине-зелёные водоросли размножаются и другими способом - образованием спор. Известно, что большинство сине-зелёных водорослей способно синтезировать все вещества своей клетки за счёт энергии света. Отдел сине-зелёных водорослей считают старейшей группой автотрофных растений на Земле (примитивное строение клетки, отсутствие полового размножения, жгутиковых стадий). По цитологии сине-зелёные водоросли сходны с бактериями.
Бурые водоросли - многоклеточные организмы сложного строения, бурого и синевато-бурого цвета. Продукт ассимиляции - полисахариды, масло. Бурые водоросли - неподвижные, прикреплённые формы. Размножение вегетативное, бесполое и половое, с чередованием гаметофита и спорофита. При половом размножении развиваются изогаметы, гетерогаметы или антерозоиды с яйцеклетками. Зооспоры и гаметы снабжены двумя жгутиками, расположенными с боков, неравной длинны и направленными в разные стороны. Бурые водоросли живут в основном в море, за исключением нескольких пресноводных видов.
Красные водоросли - многоклеточные, очень редко одноклеточные, сложного строения, красного или голубоватого цвета. У громадного большинства красных водорослей зигота не сразу прорастает в новое растение, а проходит очень сложный путь развития, прежде чем из неё образуются новые споры, прорастающие уже в новые растения. Споры собранны в компактные группы, носящие название цистокарпа, последний часто имеет специальную оболочку. Цистокарп всегда тесно связан с тем растением, на котором происходило оплодотворение яйцеклетки и дальнейшее развитие.
Тема: КЛАССИФИКАЦИЯ РАСТЕНИЙ. ВОДОРОСЛИ. План лекции 1. Классификация растений. Систематика. 2. Общая характеристика водорослей. 3. Отдел Синезеленые водоросли (Cyanophyta). 4. Отдел Желтозеленые водоросли (Xanthophyta). 5. Отдел Диатомовые водоросли (Diatomophyta). 6. Отдел Бурые водоросли (Phaeophyta). 7. Отдел Красные водоросли (Rhodophyta). 8. Отдел Зеленые водоросли (Chlorophyta). 9. Отдел Харовые водоросли (Charophyta).
 1. Классификация растений. Систематика – наука о разнообразии живых организ мов, методах их описания и закономерностях возникновения. Конечной целью систематики является построение системы органического мира – единой классификации. Перед систематикой стоят две задачи: – распределение живых существ по группам. – описание и наименование живых существ. В соответствии с этими задачами систематика делится на две состав ляющие ее дисциплины: аксономию и номенклатуру. т
1. Классификация растений. Систематика – наука о разнообразии живых организ мов, методах их описания и закономерностях возникновения. Конечной целью систематики является построение системы органического мира – единой классификации. Перед систематикой стоят две задачи: – распределение живых существ по группам. – описание и наименование живых существ. В соответствии с этими задачами систематика делится на две состав ляющие ее дисциплины: аксономию и номенклатуру. т
 Таксономия – наука об объединении живых существ в группы на основании анализа присущих им признаков. Наиболее значимые признаки называют диагностическими. На основании диагностических признаков организмы объединяются в группы, называемые таксонами. Таксон – группа индивидуумов, создаваемая в процессе классифицирования. Иерархия – расположение групп объектов в порядке соподчинения, т. е. включения одних групп в состав других. Для всех групп организмов принята единая система иерархии, образованная общепринятыми таксономическими категориями.
Таксономия – наука об объединении живых существ в группы на основании анализа присущих им признаков. Наиболее значимые признаки называют диагностическими. На основании диагностических признаков организмы объединяются в группы, называемые таксонами. Таксон – группа индивидуумов, создаваемая в процессе классифицирования. Иерархия – расположение групп объектов в порядке соподчинения, т. е. включения одних групп в состав других. Для всех групп организмов принята единая система иерархии, образованная общепринятыми таксономическими категориями.
 Таксономические категории. 1. Imperia империя; 2. Dominion доминион; 3. Regnum царство; 4. Divislo отдел, Phylum филум; Typos тип; 5. Classis класс; 6. Ordo порядок, отряд; 7. Familia семейство; 8. Tribus триба, колено; 9. Genus род; 10. Sectio секция; 11. Series ряд, серия; 12. Species вид; 13. Var. Setas разновидность, вариетет; 14. Forma форма; 15. Forma specialis специализированная форма. Приставки «под » (sub) и «над » (super), применяют для дополнительных категорий, например, subregnum – подцарство или superordo – надпорядок
Таксономические категории. 1. Imperia империя; 2. Dominion доминион; 3. Regnum царство; 4. Divislo отдел, Phylum филум; Typos тип; 5. Classis класс; 6. Ordo порядок, отряд; 7. Familia семейство; 8. Tribus триба, колено; 9. Genus род; 10. Sectio секция; 11. Series ряд, серия; 12. Species вид; 13. Var. Setas разновидность, вариетет; 14. Forma форма; 15. Forma specialis специализированная форма. Приставки «под » (sub) и «над » (super), применяют для дополнительных категорий, например, subregnum – подцарство или superordo – надпорядок
 Номенклатура – система правил описания живых существ и присвоения названий организмам и их группам с целью облегчения их идентификации. Чтобы упорядочить этот процесс и подчинить его определенным правилам, создаются номенклатурные кодексы. Номенклатурные кодексы – своды правил, регламентирующих принципы номенклатуры той или иной группы живых существ.
Номенклатура – система правил описания живых существ и присвоения названий организмам и их группам с целью облегчения их идентификации. Чтобы упорядочить этот процесс и подчинить его определенным правилам, создаются номенклатурные кодексы. Номенклатурные кодексы – своды правил, регламентирующих принципы номенклатуры той или иной группы живых существ.
 Основные номенклатурные кодексы. Ø «Международный кодекс ботанической номенклатуры» (ICBN); Ø «Международный кодекс зоологической номенклатуры (ICZN)» ; Ø «Международный кодекс номенклатуры бактерий (ICNB)» ; Ø «Международный кодекс классификации и номенклатуры вирусов» (ICVCN); Ø «Международный кодекс номенклатуры культурных растений» (ICNCP). Идет разработка единого «Международного кодекса биологической номенклатуры» (Bio. Code).
Основные номенклатурные кодексы. Ø «Международный кодекс ботанической номенклатуры» (ICBN); Ø «Международный кодекс зоологической номенклатуры (ICZN)» ; Ø «Международный кодекс номенклатуры бактерий (ICNB)» ; Ø «Международный кодекс классификации и номенклатуры вирусов» (ICVCN); Ø «Международный кодекс номенклатуры культурных растений» (ICNCP). Идет разработка единого «Международного кодекса биологической номенклатуры» (Bio. Code).
 Первый свод правил ботанической номенклатуры 1867 г. Альфонс Декандоль. В 1952 г. в Стокгольме был принят первый «Международный Кодекс ботанической номенклатуры» . Все изменения и дополнения вносятся в номенклатурные кодексы в соответствии с решениями международных конгрессов – ботанического (IBC), зоологического (IZC), бактериологического (ICB) и вирусологического (IVC). Международные форумы собираются раз в 4 – 5 лет для решения важнейших таксономических и номенклатурных проблем.
Первый свод правил ботанической номенклатуры 1867 г. Альфонс Декандоль. В 1952 г. в Стокгольме был принят первый «Международный Кодекс ботанической номенклатуры» . Все изменения и дополнения вносятся в номенклатурные кодексы в соответствии с решениями международных конгрессов – ботанического (IBC), зоологического (IZC), бактериологического (ICB) и вирусологического (IVC). Международные форумы собираются раз в 4 – 5 лет для решения важнейших таксономических и номенклатурных проблем.
 2. Общая характеристика водорослей. Водоросли (Algae) – совокупность отделов, преимущественно водных организмов. Водоросли не имеют настоящих органов и тканей, в связи с чем, тело многоклеточных форм представлено талломом. Особенности строения. Водоросли могут быть одноклеточными, колониальными и многоклеточными; в зависимости от расположения клеток – нитчатыми, пластинчатыми или сложно разветвленными.
2. Общая характеристика водорослей. Водоросли (Algae) – совокупность отделов, преимущественно водных организмов. Водоросли не имеют настоящих органов и тканей, в связи с чем, тело многоклеточных форм представлено талломом. Особенности строения. Водоросли могут быть одноклеточными, колониальными и многоклеточными; в зависимости от расположения клеток – нитчатыми, пластинчатыми или сложно разветвленными.
 Вегетативные клетки таллома водоросли состоят: Ø твердая клеточная стенка из целлюлозы и пектиновых веществ. Ø цитоплазма. вакуоль с клеточным соком. ядеро одно или несколько. хлоропласты (пластинчатые, цилиндрические, лентовидные, чашевидные, звездчатые и т. д.). üпиреноиды белковые тельца, вокруг которых откладываются крахмал, масла, лейкозин, волютин.
Вегетативные клетки таллома водоросли состоят: Ø твердая клеточная стенка из целлюлозы и пектиновых веществ. Ø цитоплазма. вакуоль с клеточным соком. ядеро одно или несколько. хлоропласты (пластинчатые, цилиндрические, лентовидные, чашевидные, звездчатые и т. д.). üпиреноиды белковые тельца, вокруг которых откладываются крахмал, масла, лейкозин, волютин.

 Вегетативное размножение у одноклеточных происходит путем деления клетки, у колониальных – распадом колонии на части, у многоклеточных – частями таллома, а иногда путем образования специальных органов вегетативного размножения.
Вегетативное размножение у одноклеточных происходит путем деления клетки, у колониальных – распадом колонии на части, у многоклеточных – частями таллома, а иногда путем образования специальных органов вегетативного размножения.
 Собственно бесполое размножение осуществляется с помощью зооспор или спор, возникающих внутри вегетативных клеток или в особых органах – зооспорангиях или спорангиях. После выхода в воду зооспоры сбрасывают жгутики, покрываются клеточной оболочкой и прорастают в новую особь.
Собственно бесполое размножение осуществляется с помощью зооспор или спор, возникающих внутри вегетативных клеток или в особых органах – зооспорангиях или спорангиях. После выхода в воду зооспоры сбрасывают жгутики, покрываются клеточной оболочкой и прорастают в новую особь.
 Половой процесс у водорослей может быть в различных формах: изо, гетеро и оогамии. Изо и гетерогаметы образуются в обычных вегетативных клетках, яйцеклетки – в оогониях, сперматозоиды – в антеридиях. У некоторых водорослей наблюдают половой процесс в виде соматогамии (конъюгации), слияние протопластов двух вегетативных клеток. Конечный результат полового процесса – образование зиготы.
Половой процесс у водорослей может быть в различных формах: изо, гетеро и оогамии. Изо и гетерогаметы образуются в обычных вегетативных клетках, яйцеклетки – в оогониях, сперматозоиды – в антеридиях. У некоторых водорослей наблюдают половой процесс в виде соматогамии (конъюгации), слияние протопластов двух вегетативных клеток. Конечный результат полового процесса – образование зиготы.
 Особи, формирующие споры называют спорофитом, а образующие гаметы – гаметофитом. Они могут быть обоеполыми и раздельнополыми. У большинства водорослей это самостоятельные растения. У некоторых видов споры и гаметы образуются на одних и тех же растениях, их называют спорогаметофитами. Спорофит и гаметофит могут иметь одинаковое строение (изоморфная смена поколений) или разное (гетероморфная смена поколений).
Особи, формирующие споры называют спорофитом, а образующие гаметы – гаметофитом. Они могут быть обоеполыми и раздельнополыми. У большинства водорослей это самостоятельные растения. У некоторых видов споры и гаметы образуются на одних и тех же растениях, их называют спорогаметофитами. Спорофит и гаметофит могут иметь одинаковое строение (изоморфная смена поколений) или разное (гетероморфная смена поколений).
 Соотношение фаз развития в жизненном цикле зависит от того, когда происходит мейоз: Ø если прорастании зиготы – тогда вся жизнь проходит в гаплоидной фазе, диплоидна лишь зигота.
Соотношение фаз развития в жизненном цикле зависит от того, когда происходит мейоз: Ø если прорастании зиготы – тогда вся жизнь проходит в гаплоидной фазе, диплоидна лишь зигота.
 Ø при образовании гамет – тогда вся жизнь проходит в диплоидной фазе, гаплоидны лишь гаметы;
Ø при образовании гамет – тогда вся жизнь проходит в диплоидной фазе, гаплоидны лишь гаметы;
 Ø если при образовании спор – тогда в жизненном цикле чередуются диплоидная и гаплоидная фазы.
Ø если при образовании спор – тогда в жизненном цикле чередуются диплоидная и гаплоидная фазы.
 Распространение и хозяйственное значение водорослей. Место обитания: Ø Вода, преимущественно морская. Часть водорослей плавающая на поверхности и в верхнем слое воды образу планктон, другая часть свободно лежат на дне или прикрепляются к нему называется бентос. Ø Водоросли живут на почве, в почве. Ø В атмосферном воздухе (некоторые виды хлореллы). Отдельные виды, попадая вместе с бактериями на бесплодные субстраты, становятся пионерами их заселения.
Распространение и хозяйственное значение водорослей. Место обитания: Ø Вода, преимущественно морская. Часть водорослей плавающая на поверхности и в верхнем слое воды образу планктон, другая часть свободно лежат на дне или прикрепляются к нему называется бентос. Ø Водоросли живут на почве, в почве. Ø В атмосферном воздухе (некоторые виды хлореллы). Отдельные виды, попадая вместе с бактериями на бесплодные субстраты, становятся пионерами их заселения.
 Ø Ø Ø Фиксируют и накапливают в почве азот. Входят в состав комплексных организмов лишайников. Служат кормом для рыб и водоплавающих птиц. Получают муку для корма с. х. животных. Используются в качестве удобрений. Бурые и красные водоросли источник агара, а их зола – сырье для получения брома и йода. Ø Некоторые водоросли используются в пищу человеком и служат сырьем для фармацевтической промышленности.
Ø Ø Ø Фиксируют и накапливают в почве азот. Входят в состав комплексных организмов лишайников. Служат кормом для рыб и водоплавающих птиц. Получают муку для корма с. х. животных. Используются в качестве удобрений. Бурые и красные водоросли источник агара, а их зола – сырье для получения брома и йода. Ø Некоторые водоросли используются в пищу человеком и служат сырьем для фармацевтической промышленности.
 3. Отдел Синезеленые водоросли (Cyanophyta). Это прокарироты – цианобактерии. Общее число видов около 1, 4 тысяч. Строение. Это колониальные и многоклеточные, реже одноклеточные организмы разнообразной окраски (сине зеленой, оливковой, темно зеленой). Окраска обусловлена пигментами, которые содержатся в клетках в разных сочетаниях: Øфикоцианином (сине зеленого цвета); Øхлорофиллом (зеленого цвета); Øкаротиноидами (желтого цвета); Øфикоэритрином (красного цвета).
3. Отдел Синезеленые водоросли (Cyanophyta). Это прокарироты – цианобактерии. Общее число видов около 1, 4 тысяч. Строение. Это колониальные и многоклеточные, реже одноклеточные организмы разнообразной окраски (сине зеленой, оливковой, темно зеленой). Окраска обусловлена пигментами, которые содержатся в клетках в разных сочетаниях: Øфикоцианином (сине зеленого цвета); Øхлорофиллом (зеленого цвета); Øкаротиноидами (желтого цвета); Øфикоэритрином (красного цвета).
 Строение В клетках сине зеленых водорослей нет обособленных ядер, хлоропластов, митохондрий и вакуолей с клеточным соком. Клеточная стенка в основном пектиновая и легко ослизняется. Полость клетки заполнена цитоплазмой, которую подразделяют на два слоя: хроматоплазму плотный постенный слой, содержащий мембраны с пигментами, и центроплазму (нуклеоид) – бесцветную центральную часть, содержащую ДНК.
Строение В клетках сине зеленых водорослей нет обособленных ядер, хлоропластов, митохондрий и вакуолей с клеточным соком. Клеточная стенка в основном пектиновая и легко ослизняется. Полость клетки заполнена цитоплазмой, которую подразделяют на два слоя: хроматоплазму плотный постенный слой, содержащий мембраны с пигментами, и центроплазму (нуклеоид) – бесцветную центральную часть, содержащую ДНК.
 Многоклеточные сине зеленые водоросли, как правило, имеют форму нитей. Протопласты соседних клеток соединены плазмодесмами. Рост нитей осуществляется за счет простого деления клеток. Среди однородных клеток, составляющих нить, имеются более крупные толстостенные клетки с желто бурым содержимым – етероцисты. г Виды синезеленых водорослей: 1 осциллатория (общий вид нити); 2 анабена (общий вид нити); 3 глеокапса; 4 хроококк (с спора, г гетероциста)
Многоклеточные сине зеленые водоросли, как правило, имеют форму нитей. Протопласты соседних клеток соединены плазмодесмами. Рост нитей осуществляется за счет простого деления клеток. Среди однородных клеток, составляющих нить, имеются более крупные толстостенные клетки с желто бурым содержимым – етероцисты. г Виды синезеленых водорослей: 1 осциллатория (общий вид нити); 2 анабена (общий вид нити); 3 глеокапса; 4 хроококк (с спора, г гетероциста)
 Питаются сине зеленые водоросли автотрофно или миксотрофно (смешанно). В качестве запасных веществ образуются: Ø гликопротеид похожий на гликоген; Ø волютин (белок); Ø цианофицин (липопротеид). Размножение преимущественно вегетативное. У одноклеточных форм путем деления клетки, у многоклеточных – путем распада нитей на части. При неблагоприятных условиях из клеток формируются толстостенные споры.
Питаются сине зеленые водоросли автотрофно или миксотрофно (смешанно). В качестве запасных веществ образуются: Ø гликопротеид похожий на гликоген; Ø волютин (белок); Ø цианофицин (липопротеид). Размножение преимущественно вегетативное. У одноклеточных форм путем деления клетки, у многоклеточных – путем распада нитей на части. При неблагоприятных условиях из клеток формируются толстостенные споры.
 Распространение и хозяйственное значение. Живут в пресных и морских водах, на почве и в почве, на голых скалах, в снегу и горячих источниках. Некоторые планктонные виды вызывают «цветение» воды в водоемах, другие очищают воду, минерализуя продукты гниения. Почвенные виды сине зеленых водорослей способны усваивать атмосферный азот. Многие виды вступают в симбиоз с грибами, образуя лишайники. Наиболее обычными в Беларуси являются представители родов анабена (Anabaena) и носток (Nostoc).
Распространение и хозяйственное значение. Живут в пресных и морских водах, на почве и в почве, на голых скалах, в снегу и горячих источниках. Некоторые планктонные виды вызывают «цветение» воды в водоемах, другие очищают воду, минерализуя продукты гниения. Почвенные виды сине зеленых водорослей способны усваивать атмосферный азот. Многие виды вступают в симбиоз с грибами, образуя лишайники. Наиболее обычными в Беларуси являются представители родов анабена (Anabaena) и носток (Nostoc).
 4. Отдел Желтозеленые водоросли (Xanthophyta). Отдел объединяет около 2, 5 тысяч видов. Желтозеленые водоросли представлены одноклеточными, колониальными, многоклеточными и неклеточными организмами. Распространение. В водоемах с пресной, реже соленой водой, где они служат важным компонентом планктона, а иногда и бентоса, живут также в почве, на камнях. Строение. Стенки их клеток чаще пектиновые, реже целлюлозные. Ядро у большинства видов одно, реже – ядер много. Хлоропласты, кроме хлорофилла, содержат каротиноиды, придающие таллому желто зеленую окраску. Пиреноиды встречаются редко. Запасные продукты. Масло, иногда в виде лейкозина и волютина.
4. Отдел Желтозеленые водоросли (Xanthophyta). Отдел объединяет около 2, 5 тысяч видов. Желтозеленые водоросли представлены одноклеточными, колониальными, многоклеточными и неклеточными организмами. Распространение. В водоемах с пресной, реже соленой водой, где они служат важным компонентом планктона, а иногда и бентоса, живут также в почве, на камнях. Строение. Стенки их клеток чаще пектиновые, реже целлюлозные. Ядро у большинства видов одно, реже – ядер много. Хлоропласты, кроме хлорофилла, содержат каротиноиды, придающие таллому желто зеленую окраску. Пиреноиды встречаются редко. Запасные продукты. Масло, иногда в виде лейкозина и волютина.
 Вегетативное размножение осуществляется путем деления клеток или частями таллома. Половое размножение известно у немногих видов. Половой процесс преимущественно изогамный, изредка оогамный. Одноклеточные особи, а также ооспоры и сперматозоиды имеют два жгутика неодинаковой длины: один из них короткий, прямой, гладкий, а другой – длинный, перистый. Ботридиум: 1 слоевище; 2 выход зооспор; 3 зооспора
Вегетативное размножение осуществляется путем деления клеток или частями таллома. Половое размножение известно у немногих видов. Половой процесс преимущественно изогамный, изредка оогамный. Одноклеточные особи, а также ооспоры и сперматозоиды имеют два жгутика неодинаковой длины: один из них короткий, прямой, гладкий, а другой – длинный, перистый. Ботридиум: 1 слоевище; 2 выход зооспор; 3 зооспора
 Представителями отдела являются виды рода вошерия (Vaucheria). Распространение. Обитают в пресной стоячей и проточной воде, а также по берегам высыхающих водоемов, прикрепляясь к почве с помощью ризоидов. Строение. Таллом ветвистый, многоядерный, но между ядрами нет клеточных перегородок. В цитоплазме имеются многочисленные мелкие хлоропласты дисковидной формы.
Представителями отдела являются виды рода вошерия (Vaucheria). Распространение. Обитают в пресной стоячей и проточной воде, а также по берегам высыхающих водоемов, прикрепляясь к почве с помощью ризоидов. Строение. Таллом ветвистый, многоядерный, но между ядрами нет клеточных перегородок. В цитоплазме имеются многочисленные мелкие хлоропласты дисковидной формы.
 Размножение. При бесполом размножении образуется зооспорангий с одной большой зооспорой, снабженной многочисленными жгутиками. Зооспора некоторое время плавает в воде, а затем сбрасывает жгутики и прорастает в новый таллом. Половой процесс оогамный. Антеридии и оогонии одноклеточные. После периода покоя зигота делится путем мейоза и прорастает, образуя новую особь. Вошерия: 1 – 2 – этапы образование зооспорангия; 3 – 4 выход зооспоры; 5 участок зооспоры при большом увеличении; 6 – сперматозоид; 7 фрагмент слоевища вошерии; 8 участок слоевища (оогонии, антеридии).
Размножение. При бесполом размножении образуется зооспорангий с одной большой зооспорой, снабженной многочисленными жгутиками. Зооспора некоторое время плавает в воде, а затем сбрасывает жгутики и прорастает в новый таллом. Половой процесс оогамный. Антеридии и оогонии одноклеточные. После периода покоя зигота делится путем мейоза и прорастает, образуя новую особь. Вошерия: 1 – 2 – этапы образование зооспорангия; 3 – 4 выход зооспоры; 5 участок зооспоры при большом увеличении; 6 – сперматозоид; 7 фрагмент слоевища вошерии; 8 участок слоевища (оогонии, антеридии).
 5. Отдел диатомовые водоросли (Diatomophyta). Отдел объединяет более 10 тысяч видов. Это одноклеточные организмы, иногда объединяющиеся в колонии. Распространение. Повсеместно. Они обитают в соленых и пресных водоемах, на влажной почве, скалах, стволах деревьев. Строение. Клеточные стенки состоят из кремнезема (Si. O 2), образующего панцирь. Створки теки, верхняя – эпитека, нижняя – гипотека. В створках имеются сквозные отверстия поры, а также пустоты. Скульптура поверхности створок очень разнообразна и имеет значение при определении видов. Внутри клетки расположены протопласт и вакуоли. Ядро одно. Хлоропласты имеют бурую окраску, так как хлорофилл в них замаскирован пигментами – каротиноидами и диатомином. Запасные продукты откладываются в виде жирных масел, волютина и лейкозина.
5. Отдел диатомовые водоросли (Diatomophyta). Отдел объединяет более 10 тысяч видов. Это одноклеточные организмы, иногда объединяющиеся в колонии. Распространение. Повсеместно. Они обитают в соленых и пресных водоемах, на влажной почве, скалах, стволах деревьев. Строение. Клеточные стенки состоят из кремнезема (Si. O 2), образующего панцирь. Створки теки, верхняя – эпитека, нижняя – гипотека. В створках имеются сквозные отверстия поры, а также пустоты. Скульптура поверхности створок очень разнообразна и имеет значение при определении видов. Внутри клетки расположены протопласт и вакуоли. Ядро одно. Хлоропласты имеют бурую окраску, так как хлорофилл в них замаскирован пигментами – каротиноидами и диатомином. Запасные продукты откладываются в виде жирных масел, волютина и лейкозина.
 Диатомовые водоросли: А пиннулярия: 1, 2 панцирь (вид со стороны створки и пояска); 3 клетка (вид со стороны створки); У узелки; Ш шов; С створки; П пояски; Э эпитека; Г гипотека.
Диатомовые водоросли: А пиннулярия: 1, 2 панцирь (вид со стороны створки и пояска); 3 клетка (вид со стороны створки); У узелки; Ш шов; С створки; П пояски; Э эпитека; Г гипотека.
 Вегетативное размножение осуществляется митотическим делением протопласта. Затем дочерние протопласты расходятся, неся по одной из створок. В последствии каждая из дочерних особей достраивает новую створку. Серия таких делений ведет к последовательному измельчению особей. В связи с этим половой процесс приводит не к численному увеличению особей, а к восстановлению их нормальных размеров. Вегетативное размножение пиннулярии: 1 – эпитека, 2 – гипотека, 10 – створка, 11 – поясок
Вегетативное размножение осуществляется митотическим делением протопласта. Затем дочерние протопласты расходятся, неся по одной из створок. В последствии каждая из дочерних особей достраивает новую створку. Серия таких делений ведет к последовательному измельчению особей. В связи с этим половой процесс приводит не к численному увеличению особей, а к восстановлению их нормальных размеров. Вегетативное размножение пиннулярии: 1 – эпитека, 2 – гипотека, 10 – створка, 11 – поясок
 Форма полового процесса очень своеобразна: измельченные особи приближаются одна к другой, сбрасывают теки и покрываются слизью. Каждая из клеток делится путем мейоза, в результате образуются четыре гаплоидные клетки – тетрада. Две или три клетки у каждой особи отмирают, а остальные сливаются попарно. Зиготу называют ауксоспорой. Из нее вырастает новая особь нормальных размеров.
Форма полового процесса очень своеобразна: измельченные особи приближаются одна к другой, сбрасывают теки и покрываются слизью. Каждая из клеток делится путем мейоза, в результате образуются четыре гаплоидные клетки – тетрада. Две или три клетки у каждой особи отмирают, а остальные сливаются попарно. Зиготу называют ауксоспорой. Из нее вырастает новая особь нормальных размеров.
 Использование диатомовых водорослей: Ø Из створок отмерших диатомовых водорослей образовались мощные отложения горных пород – диатомита и трепела. Их используют при производстве динамита, материалов для звуковой и тепловой изоляции, при шлифовке металлов, изготовлении фильтров. Ø Участвуют в естественных процессах очищения воды. Ø Соединения кремния диатомовых перспективны для использования в нанотехнологии, для получения материалов с заранее заданными свойствами. Наиболее массовыми в пресных водоемах Республики Беларусь являются виды родов пиннулария, табеллария и циклотелла.
Использование диатомовых водорослей: Ø Из створок отмерших диатомовых водорослей образовались мощные отложения горных пород – диатомита и трепела. Их используют при производстве динамита, материалов для звуковой и тепловой изоляции, при шлифовке металлов, изготовлении фильтров. Ø Участвуют в естественных процессах очищения воды. Ø Соединения кремния диатомовых перспективны для использования в нанотехнологии, для получения материалов с заранее заданными свойствами. Наиболее массовыми в пресных водоемах Республики Беларусь являются виды родов пиннулария, табеллария и циклотелла.
 6. Отдел Бурые водоросли (Phaeophyta). Общее число видов около 1, 5 тысяч. Распространение. Главным образом в морях и океанах всего мира. Окраска таллома от оливково желтоватой до темно бурой. Цвет обусловлен смесью разных пигментов: хлорофилла, каротиноидов, фукоксантина (бурого цвета).
6. Отдел Бурые водоросли (Phaeophyta). Общее число видов около 1, 5 тысяч. Распространение. Главным образом в морях и океанах всего мира. Окраска таллома от оливково желтоватой до темно бурой. Цвет обусловлен смесью разных пигментов: хлорофилла, каротиноидов, фукоксантина (бурого цвета).
 Строение. Таллом бурых водорослей многоклеточный. Среди них есть гиганты, достигающие иногда 30 50 м. У высокоорганизованных видов формируя обособленные комплексы клеток, напоминающие ассимиляционную, запасающую, механическую и проводящую ткани. Такая дифференциация обусловлена расчленением таллома на участки, выполняющие различные функции: ризоиды, осевую («стеблевую») часть и филлоиды («листовую» часть). Клетки бурых водорослей одноядерные. Многочисленные хлоропласты чаще
Строение. Таллом бурых водорослей многоклеточный. Среди них есть гиганты, достигающие иногда 30 50 м. У высокоорганизованных видов формируя обособленные комплексы клеток, напоминающие ассимиляционную, запасающую, механическую и проводящую ткани. Такая дифференциация обусловлена расчленением таллома на участки, выполняющие различные функции: ризоиды, осевую («стеблевую») часть и филлоиды («листовую» часть). Клетки бурых водорослей одноядерные. Многочисленные хлоропласты чаще
 Запасные продукты накапливаются в виде ламинарина, маннита и жирных масел. Пектиново целлюлозные клеточные стенки легко ослизняются. Рост таллома верхушечный или интеркалярный. Продолжительность жизни достигает нескольких лет. Вегетативное размножение может осуществляться участками таллома. Бесполое размножение происходит при помощи многочисленных двухжгутиковых зооспор, образующихся в одногнездных, изредка многогнездных зооспорангиях, или неподвижных тетраспор, образующихся в одногнездных тетраспорангиях. Половой процесс. Изогамный, гетерогамный и оогамный.
Запасные продукты накапливаются в виде ламинарина, маннита и жирных масел. Пектиново целлюлозные клеточные стенки легко ослизняются. Рост таллома верхушечный или интеркалярный. Продолжительность жизни достигает нескольких лет. Вегетативное размножение может осуществляться участками таллома. Бесполое размножение происходит при помощи многочисленных двухжгутиковых зооспор, образующихся в одногнездных, изредка многогнездных зооспорангиях, или неподвижных тетраспор, образующихся в одногнездных тетраспорангиях. Половой процесс. Изогамный, гетерогамный и оогамный.
 У всех бурых водорослей, кроме фукусовых, в жизненном цикле хорошо выражена смена ядерных фаз. Мейоз происходит в зооспорангиях или тетраспорангиях. Зооспоры или тетраспоры дают начало гаметофиту (n), который бывает обоеполым или раздельнополым. Зигота без периода покоя прорастает в спорофит (2 n). У видов разных родов характер смены ядерных фаз различный: у одних спорофит и гаметофит по внешнему виду не различаются, у других спорофит более мощный и более долговечный, чем гаметофит.
У всех бурых водорослей, кроме фукусовых, в жизненном цикле хорошо выражена смена ядерных фаз. Мейоз происходит в зооспорангиях или тетраспорангиях. Зооспоры или тетраспоры дают начало гаметофиту (n), который бывает обоеполым или раздельнополым. Зигота без периода покоя прорастает в спорофит (2 n). У видов разных родов характер смены ядерных фаз различный: у одних спорофит и гаметофит по внешнему виду не различаются, у других спорофит более мощный и более долговечный, чем гаметофит.
 Бурые водоросли имеют большое значение как кормовые, пищевые, лекарственные и технические растения. Наибольшее значение в качестве пищевого сырья имеют виды рода ламинария (Laminaria). Виды рода – многолетние растения, которые растут в Охотском, Белом, Черном, Красном и Японском морях, имеют длинное листовидное слоевище зеленовато коричневого цвета. Нижняя часть таллома своим внешний видом похожа на черешок листа. С помощью ризоидов ламинария прикрепляется к камням.
Бурые водоросли имеют большое значение как кормовые, пищевые, лекарственные и технические растения. Наибольшее значение в качестве пищевого сырья имеют виды рода ламинария (Laminaria). Виды рода – многолетние растения, которые растут в Охотском, Белом, Черном, Красном и Японском морях, имеют длинное листовидное слоевище зеленовато коричневого цвета. Нижняя часть таллома своим внешний видом похожа на черешок листа. С помощью ризоидов ламинария прикрепляется к камням.
 7. Отдел Красные водоросли (Rhodophyta). Общее число видов 4 тысячи. Распространены в морях тропических и субтропических областей, иногда в водоемах территорий с умеренным климатом и немногие являются пресноводными или аэрофитами (поселяются на почве и коре деревьев). Строение. Их таллом имеет вид кустиков, составленных из многоклеточных ветвящихся нитей, реже пластинчатый или листовидный, иногда длиной до 2 м. Имеются признаки тканеподобной дифференциации. Рост диффузный (специализированных зон роста нет) или верхушечный. Подвижные формы в жизненном цикле отсутствуют.
7. Отдел Красные водоросли (Rhodophyta). Общее число видов 4 тысячи. Распространены в морях тропических и субтропических областей, иногда в водоемах территорий с умеренным климатом и немногие являются пресноводными или аэрофитами (поселяются на почве и коре деревьев). Строение. Их таллом имеет вид кустиков, составленных из многоклеточных ветвящихся нитей, реже пластинчатый или листовидный, иногда длиной до 2 м. Имеются признаки тканеподобной дифференциации. Рост диффузный (специализированных зон роста нет) или верхушечный. Подвижные формы в жизненном цикле отсутствуют.
 Окраска разнообразная обусловлена различными пигментами: хлорофилл, каротиноидов, фикоэритрина, фикоцианина. Хлоропласты чаще в форме дисков без пиреноидов. Запасные вещества в виде багрянкового крахмала. Пектиново целлюлозные клеточные стенки вместе с межклеточным веществом у некоторых видов ослизняются настолько сильно, что весь таллом приобретает слизистую консистенцию. У других видов клеточные стенки инкрустированы известью, придающей таллому жесткость.
Окраска разнообразная обусловлена различными пигментами: хлорофилл, каротиноидов, фикоэритрина, фикоцианина. Хлоропласты чаще в форме дисков без пиреноидов. Запасные вещества в виде багрянкового крахмала. Пектиново целлюлозные клеточные стенки вместе с межклеточным веществом у некоторых видов ослизняются настолько сильно, что весь таллом приобретает слизистую консистенцию. У других видов клеточные стенки инкрустированы известью, придающей таллому жесткость.
 Вегетативное размножение происходит посредством образования дополнительных побегов, которые берут начало от подошвы или от стелющихся ветвей. Вертикальное слоевище в определенный период отмирает и остается только базальная часть растения, которая через какое то время прорастает, образуя новые побеги. Самые примитивные багрянки размножаются только вегетативно. Для бесполого размножения служат различного рода споры. У низкоорганизованных багрянок бесполое размножение осуществляется моноспорами. Моноспоры не имеют жгутика и оболочки, после выхода из материнской клетки они способны к амебоидному движению. У одних видов моноспоры образуются в любой клетке таллома и до своего выхода не отличаются от вегетативных клеток, у других – моноспорангии образуются на веточках ограниченного роста.
Вегетативное размножение происходит посредством образования дополнительных побегов, которые берут начало от подошвы или от стелющихся ветвей. Вертикальное слоевище в определенный период отмирает и остается только базальная часть растения, которая через какое то время прорастает, образуя новые побеги. Самые примитивные багрянки размножаются только вегетативно. Для бесполого размножения служат различного рода споры. У низкоорганизованных багрянок бесполое размножение осуществляется моноспорами. Моноспоры не имеют жгутика и оболочки, после выхода из материнской клетки они способны к амебоидному движению. У одних видов моноспоры образуются в любой клетке таллома и до своего выхода не отличаются от вегетативных клеток, у других – моноспорангии образуются на веточках ограниченного роста.
 Половое размножение красных водорослей осуществляется через образование сложно устро енных половых органов. Женский половой орган, называемый карпогоном, возникает на концах боковых веточек. Нижняя часть его расширена, а верхняя сужена в так называемую трихогину. В нижней части кар погона находится яйцеклетка. Мужские органы – антеридии – собраны группами на концах сильно ветвящихся нитей. В них развивается по одному не подвижному сперматозоиду, называемому пермацием. с
Половое размножение красных водорослей осуществляется через образование сложно устро енных половых органов. Женский половой орган, называемый карпогоном, возникает на концах боковых веточек. Нижняя часть его расширена, а верхняя сужена в так называемую трихогину. В нижней части кар погона находится яйцеклетка. Мужские органы – антеридии – собраны группами на концах сильно ветвящихся нитей. В них развивается по одному не подвижному сперматозоиду, называемому пермацием. с

 Значение. Сырье для получения агара и йода, их используют как корм для животных, употребляют в пищу. В Беларуси встречается один из видов пресноводных водорослей – батрахоспермум четковидный Вatrachospermum (moniliforme).
Значение. Сырье для получения агара и йода, их используют как корм для животных, употребляют в пищу. В Беларуси встречается один из видов пресноводных водорослей – батрахоспермум четковидный Вatrachospermum (moniliforme).
 Морскими представителями красных водорослей являются виды родов порфира (Porphyra) и фикодрис (Phicodrys). Порфира имеют таллом в иде в однослойной или двухслойной пластинки, сужающейся в нижней части в небольшой стебелёк. Стебелёк переходит в подошву, образованную ризоидами. Растения достигают в длину метр и более. Окраска пластинок розово красная.
Морскими представителями красных водорослей являются виды родов порфира (Porphyra) и фикодрис (Phicodrys). Порфира имеют таллом в иде в однослойной или двухслойной пластинки, сужающейся в нижней части в небольшой стебелёк. Стебелёк переходит в подошву, образованную ризоидами. Растения достигают в длину метр и более. Окраска пластинок розово красная.
 Представители рода фикодрис имеют талломы до 20 см высотой, часто с сильноразветвленными стебельками. Пластины одиночные или многочисленные, кожистые, с короткоклиновидным или сердцевидным основанием.
Представители рода фикодрис имеют талломы до 20 см высотой, часто с сильноразветвленными стебельками. Пластины одиночные или многочисленные, кожистые, с короткоклиновидным или сердцевидным основанием.
 8. Отдел Зеленые водоросли (Chlorophyta). Общее число видов около 15 тысяч. Распространены повсеместно, преимущественно в пресных водоемах, некоторые в морях и очень немногие в условиях периодического увлажнения на почве, стволах деревьев, заборах, цветочных горшках. Строение. Отдел включает одноклеточные и многоклеточные формы. Органы движения у подвижных форм два, реже четыре жгутика одинаковой длины и формы. Клетки одноядерные, но могут быть и многоядерными. Хлоропласты в большинстве случаев с пиреноидами, разнообразные по форме, величине и по числу в клетке. Пигменты – хлорофилл, каротиноиды. Запасные продукты – крахмал и жирные масла. Размножение вегетативное, бесполое и половое. Половой процесс известен почти у всех видов и отличается большим разнообразием: изогамия, гетерогамия, оогамия, соматогамия.
8. Отдел Зеленые водоросли (Chlorophyta). Общее число видов около 15 тысяч. Распространены повсеместно, преимущественно в пресных водоемах, некоторые в морях и очень немногие в условиях периодического увлажнения на почве, стволах деревьев, заборах, цветочных горшках. Строение. Отдел включает одноклеточные и многоклеточные формы. Органы движения у подвижных форм два, реже четыре жгутика одинаковой длины и формы. Клетки одноядерные, но могут быть и многоядерными. Хлоропласты в большинстве случаев с пиреноидами, разнообразные по форме, величине и по числу в клетке. Пигменты – хлорофилл, каротиноиды. Запасные продукты – крахмал и жирные масла. Размножение вегетативное, бесполое и половое. Половой процесс известен почти у всех видов и отличается большим разнообразием: изогамия, гетерогамия, оогамия, соматогамия.
 Типичным представителем многоклеточных зеленых водорослей являются виды рода спирогира (Spirogyra). Место обитания. Пресные водоемы, реки, пруды, озера и торфяные болота. Строение. Нитчатый таллом состоит из одного ряда клеток. Хлоропласты по 1 2 в клетке расположены в постенном слое цитоплазмы. Они имеют вид спирально закрученных лент. Ядро находится в центре клетки и погружено в цитоплазму, тончайшие нити которой тянутся к ее постенному слою. Вакуолей несколько. Нити растут в длину за счет деления клеток.
Типичным представителем многоклеточных зеленых водорослей являются виды рода спирогира (Spirogyra). Место обитания. Пресные водоемы, реки, пруды, озера и торфяные болота. Строение. Нитчатый таллом состоит из одного ряда клеток. Хлоропласты по 1 2 в клетке расположены в постенном слое цитоплазмы. Они имеют вид спирально закрученных лент. Ядро находится в центре клетки и погружено в цитоплазму, тончайшие нити которой тянутся к ее постенному слою. Вакуолей несколько. Нити растут в длину за счет деления клеток.
 Вегетативное размножение. Частями таллома. Половой процесс. Конъюгация. Две особи располагаются параллельно. В их клетках возникают выпячивания стенок, растущие навстречу другу. В месте стыка стенки клеток ослизняются, образуя конъюгационный канал, по которому протопласт из клетки одной особи переходит в клетку другой особи. Образуется крупная шаровидная зигота с толстой стенкой. Зигота делится мейозом. Образуются четыре гаплоидные клетки, три из них отмирают, а одна прорастает в новую особь.
Вегетативное размножение. Частями таллома. Половой процесс. Конъюгация. Две особи располагаются параллельно. В их клетках возникают выпячивания стенок, растущие навстречу другу. В месте стыка стенки клеток ослизняются, образуя конъюгационный канал, по которому протопласт из клетки одной особи переходит в клетку другой особи. Образуется крупная шаровидная зигота с толстой стенкой. Зигота делится мейозом. Образуются четыре гаплоидные клетки, три из них отмирают, а одна прорастает в новую особь.
 9. Отдел Харовые водоросли (Charophyta). Отдел объединяет не более 700 видов. Строение. Это макроскопические водоросли, внешне сходные с некоторыми высшими растениями. Высота их таллома составляет обычно 20 30 см, но может достигать и 1 2 м. Боковые ветви имеют ограниченный рост, расположены мутовками в многоклеточных узлах. Междоузлия состоят из одной длинной клетки, которая может обрастать корой из узких клеток. Оболочки клеток пропитаны известью. Хлоропласты зелёные, содержат хлорофиллы и ликопин. Запасное вещество – крахмал. Размножение. Вегетативное размножение осуществляется посредством специальных клубеньков на ризоидах или звездообразных скоплений клеток на нижних стеблевых узлах, которые дают начало новому таллому. Бесполое размножение отсутствует.
9. Отдел Харовые водоросли (Charophyta). Отдел объединяет не более 700 видов. Строение. Это макроскопические водоросли, внешне сходные с некоторыми высшими растениями. Высота их таллома составляет обычно 20 30 см, но может достигать и 1 2 м. Боковые ветви имеют ограниченный рост, расположены мутовками в многоклеточных узлах. Междоузлия состоят из одной длинной клетки, которая может обрастать корой из узких клеток. Оболочки клеток пропитаны известью. Хлоропласты зелёные, содержат хлорофиллы и ликопин. Запасное вещество – крахмал. Размножение. Вегетативное размножение осуществляется посредством специальных клубеньков на ризоидах или звездообразных скоплений клеток на нижних стеблевых узлах, которые дают начало новому таллому. Бесполое размножение отсутствует.









3.2. Водоросли (Algae)
3.2.1. Основные признаки и систематика водорослей
Водоросли - огромная группа растений, имеющих большое биологическое значение и очень важных для человечества (разд. 3.2.8). Они являются самыми примитивными из растений, и у них нет разделения тела на стебель, корень и листья. Поэтому первоначально их объединяли вместе с грибами в отдел Thallophyta (см. примечание на с. 43). Однако после новых научных открытий стало ясно, что водоросли ничуть не менее разнообразны, чем все остальные группы растений, вместе взятые, и что у них очень мало общих признаков. Вероятно, лучше всего считать водорослями все фотосинтезирующие организмы, выделяющие кислород, которые эволюционировали в водной среде и полностью освоили ее. Правда, некоторые водоросли вышли и на сушу, но в масштабах планеты продуктивность прибрежных и наземных форм ничтожна в сравнении с продуктивностью океанических и пресноводных водорослей. Если придерживаться такой точки зрения, то в группу водорослей следует включить и сине-зеленые водоросли (Cyanophyta). Однако, поскольку эти водоросли - прокариоты, было предложено называть их цианобактериями (Cyanobacteria), чтобы как-то отличить от эукариотических водорослей. При этом из виду упускают один очень важный факт, а именно то, что сине-зеленые водоросли при фотосинтезе выделяют кислород, а все остальные фотосинтезирующие прокариоты - нет. Для того чтобы вода могла расщепиться на водород и кислород, необходимо наличие хлорофилла и фотосистемы II (разд. 9.4.2), что является важным преимуществом перед фотосинтезирующими бактериями. О том, как было достигнуто такое преимущество, известно очень мало, хотя и обнаружены некоторые формы, занимающие промежуточное положение между сине-зелеными водорослями и бактериями. Такое истолкование связи между сине-зелеными водорослями и другими растениями, в том числе и остальными водорослями, подкрепляют данные, свидетельствующие в пользу симбиотической теории, согласно которой хлоропласты растений произошли из сине-зеленых водорослей (разд. 9.3.1).
Подводя итог, можно сказать, что термин "водоросль" сам по себе удобен, но его применение в систематике вносит ненужные осложнения. Сине-зеленые водоросли следует относить к прокариотам, а все остальные водоросли - к эукариотам.
К счастью, эукариотические водоросли вполне естественно распадаются на хорошо различимые группы, причем основным отличительным признаком является набор фотосинтетических пигментов. В современной систематике такие группы получили статус отделов. Родственные связи между отделами до сих пор не выяснены, а этот вопрос очень важен, чтобы понять происхождение высших растений и связь между прокариотами и эукариотами.
Все отделы перечислены на рис. 3.11, а на рис. 3.12 даны современные представления о том, какие связи существуют между этими отделами. Основные признаки водорослей и некоторых главных отделов приведены в табл. 3.4.

3.2.2. Бесполое размножение водорослей
У водорослей наблюдается как бесполое, так и половое размножение. Ниже кратко перечислены основные типы бесполого размножения от самого простого до самого сложного.
Вегетативное размножение . У некоторых колониальных форм колонии могут дробиться на отдельные фрагменты, которые дают начало новым более мелким колониям. У более крупных водорослей, например у Fucus, на главном талломе могут образовываться дополнительные талломы, которые отламываются и образуют новые организмы.
Фрагментация . Это явление наблюдается у нитчатых водорослей, таких, как сине-зеленые водоросли и Spirogyra. Нить расщепляется строго определенным образом вдоль, и образуются две новые нити. Это явление можно рассматривать как одну из форм вегетативного размножения.
Бинарное деление . В этом случае одноклеточный организм делится на две одинаковые половины, при этом ядро делится митотически. Продольное деление такого типа наблюдается у Euglena.
Зооспоры . Это подвижные споры, имеющие жгутики. Они образуются у многих водорослей, например у Chlamydomonas, и у некоторых грибов (см. Oomycota, табл. 3.2).
Апланоспоры . Эти неподвижные споры образуются, например, у некоторых бурых водорослей.
3.2.3. Половое размножениеводорослей
При половом размножении объединяется генетический материал двух отдельных особей одного и того же вида. Самый простой способ такого размножения у водорослей; он заключается в слиянии двух морфологически (т. е. структурно) идентичных гамет. Такой процесс называется изогамией , а гаметы - изогаметами . Изогамны Spirogyra и некоторые виды Chlamydomonas.
Если одна из гамет менее подвижна или крупней, чем другая, то такой процесс называют анизогамией . У Spirogyra гаметы не отличаются по строению, но одна из них двигается, а другая неподвижна. Это можно рассматривать как физиологическую анизогамию. Существует еще один вариант, когда одна гамета большая и неподвижная, а вторая небольшая и подвижная. Такие гаметы называются женскими и мужскими, а сам процесс называется оогамией . Оогамны Fucus и некоторые виды Chlamydomonas. Женские гаметы крупные потому, что в них находится запас питательных веществ, необходимых для развития зиготы после оплодотворения.
Все три типа полового размножения соответствуют увеличению сложности строения тела, и поэтому оогамия, хотя и встречается у некоторых простых водорослей, таких, как Chlamydomonas, в целом более распространена у более сложных водорослей, например у представителей Phaeophyta. Оогамия - это единственный способ полового размножения у растений, более высоко организованных, чем водоросли.
К сожалению, терминология, применяемая для описания гамет и органов полового размножения растений, очень запутана, особенно у водорослей. Ниже мы объясним только основные термины.
У грибов и низших растений (водорослей, мохообразных и папоротникообразных) гаметы образуются в особых структурах, которые называют гаметангиями . Мужской гаметангий называется антеридием, а женский - оогонием или архегонием.
Оогоний * - это простой женский гаметангий, который встречается у многих водорослей и грибов, а женские гаметы или гаметы, которые находятся в нем, называют оосферами . Оплодотворенная оосфера называется ооспорой ; она превращается в толстостенную покоящуюся спору, способную переживать неблагоприятные условия. Общее название для женской гаметы - яйцо или яйцеклетка , хотя иногда для обозначения яйцеклетки используют термин "оосфера"; однако это не совсем точно.
* (Оогониями называют также клетки яичников, из которых образуются ооциты у животных (см. гл. 20). )
Архегоний - это более сложный женский гаметангий, который характерен для мохообразных, папоротникообразных и многих голосеменных; архегоний будет описан далее в этой главе.
В антеридии образуются мужские гаметы, которые называются антерозоидами или сперматозоидами . Они подвижны, потому что снабжены одним или несколькими жгутиками. Такие гаметы характерны для грибов, водорослей, мохообразных, папоротникообразных и некоторых голосеменных. У животных мужские гаметы называются сперматозоидами или спермиями . Перечисленные названия приведены на рис. 3.13.

Для тех целей, которые стоят перед нами в этой главе, не столь важно, как называть разные гаметы одного и того же пола, поэтому вполне достаточно различать сперматозоиды, т. е. все мужские гаметы, и яйцеклетки, т. е. все женские гаметы.
Как и у грибов, у некоторых водорослей наблюдается гетероталличность (разд. 3.1.3).
3.2.4. Отдел Chlorophyta
Основные свойства Chlorophyta перечислены в табл. 3.4.

Таблица 3.4. Систематика и основные признаки некоторых главных групп водорослей 1)
1) (Звездочкой отмечен систематический признак. )
Chlamydomonas (хламидомонада) - одноклеточная подвижная водоросль, которая живет главным образом в стоячей воде, т. е. в прудах и канавах, особенно если вода еще и обогащена растворимыми азотистыми соединениями, например стоками со скотных дворов. Клетки этой водоросли часто встречаются в таком огромном количестве, что вода становится зеленой. Некоторые виды живут в морской воде или в солоноватых лиманах.
Строение
Хламидомонада совсем не похожа на растение, так как она активно движется и у нее имеются пульсирующие вакуоли. Строение хламидомонады изображено на рис. 3.14. На электронной микрофотографии видны типичные для эукариот органеллы: аппарат Гольджи, митохондрии, рибосомы и мелкие вакуоли. В хлоропластах многих водорослей выявлена особая структура - пиреноид . Это - белковое образование, состоящее главным образом из рибулозобисфосфаткарбоксилазы - фермента, который осуществляет фиксацию двуокиси углерода. Пиреноид участвует в запасании углеводов, например крахмала. Красный глазок воспринимает изменения в интенсивности освещения, и клетка либо перемещается туда, где интенсивность света оптимальна для фотосинтеза, либо остается на месте, если освещенность достаточна. Такая ответная реакция на свет называется фототаксисом (разд. 15.1.2). Клетка хламидомонады передвигается за счет биения двух жгутиков и ввинчивается в воду, как штопор, вращаясь вокруг продольной оси.

Рис. 3.14. A. Chlamydomonas в световом микроскопе; х 600. Б. Схема строения Chlamydomonas. В. Электронная микрофотография Chamydomonas reinhardtii. × 1400
Жизненный цикл
Жизненный цикл Chlomydomonas изображен на рис. 3.15. Взрослая особь гаплоидна.

Бесполое размножение
Бесполое размножение осуществляется с помощью зооспор. Родительская клетка теряет жгутики, и протопласт клетки делится на два-четыре дочерних протопласта (обычно на четыре). В это же время происходит митотическое деление ядра; кроме того, делится и хлоропласт. У дочерних протопластов образуются новые клеточные стенки, новые глазки и новые жгутики. В образовании новых жгутиков участвуют центриоли (базальные тельца). Клеточная стенка родительской клетки ослизняется, и дочерние клетки, которые теперь называют зооспорами, выходят наружу. Из каждой зооспоры вырастает полноценная взрослая клетка Chlamydomonas. Этот процесс изображен на рис. 3.16, А.

Половое размножение
Одни виды Chlomydomonas - гомоталличны, другие - гетероталличны; при этом разные виды могут быть изогамными, анизогамными или оогамными. Размножение изогамных видов изображено на рис. 3.16, Б. При прорастании ядро зиготы первый раз делится мейотически, при этом восстанавливается гаплоидное состояние, свойственное взрослым организмам. Высвободившиеся молодые клетки Chlomydomonas можно называть зооспорами, пока они полностью не созреют.
В прудах и других водоемах с непроточной, но чистой водой живет еще одна водоросль - неветвящаяся нитчатая водоросль Spirogyra. Большая часть видов спирогиры - плавающие формы, а нити у нее слизистые и скользкие.
Строение
Цилиндрические клетки спирогиры соединены торец в торец и образуют нить, изображенную на рис. 3.17. Все клетки идентичны, и между ними не наблюдается разделения функций. Тонкий слой цитоплазмы лежит по периферии клетки, а большая вакуоль как бы обмотана тяжами цитоплазмы. Такие тяжи удерживают ядро в центре клетки. Один или несколько спиралевидных хлоропластов лежат в тонком постенном слое цитоплазмы.

Рост и размножение
Нити спирогиры растут интеркалярно, т. е. за счет деления любой из клеток, входящих в состав нити, независимо от того, где находится эта клетка. У большинства же растений зона роста ограничена верхушечной областью. Ядро клетки спирогиры делится митотически, затем из выростов боковых стенок образуется новая поперечная клеточная стенка. Получаются две дочерние клетки, которые вырастают до нормальных размеров, в результате чего вся нить увеличивается в длину.
Как мы уже отмечали (разд. 3.2.1), бесполое размножение происходит путем фрагментации.
Половое размножение осуществляется весьма специфическим способом, характерным для нитчатых водорослей: две нити располагаются бок о бок и супротивные клетки обеих нитей соединяются короткими трубчатыми выростами. Все содержимое клетки ведет себя как гамета; процесс этот можно рассматривать как анизогамный, поскольку, хотя обе гаметы и идентичны морфологически, только одна из них подвижна и перетекает в другую клетку через соединительную трубку. Такой процесс называется конъюгацией.
3.2.5. Отдел Phaeophyta
Основные признаки Phaeophyta перечислены в табл. 3.4.
У скалистых берегов Британского побережья часто встречаются разные водоросли из рода Fucus. Они очень хорошо приспособились к достаточно суровым условиям литоральной зоны, т. е. той зоны, которая попеременно то обнажается при отливе, то снова покрывается водой.
Наиболее известны три вида Fucus, которые чаще других встречаются у побережья в трех разных зонах на разной глубине; такое явление называется зональным распределением . Эти водоросли распределяются по зонам в соответствии с их способностью выдерживать пребывание на воздухе. Перечислим основные признаки, по которым их можно узнать, и места на берегу, где их можно найти:
F. spiralis (эти плоские водоросли выбрасывает на берег море) - у высшей точки прилива. В погруженном состоянии таллом слегка закручен в спираль.
F. serratus (то, что называют обыкновенными, зубчатыми или пильчатыми водорослями) - в средней приливной зоне. Края таллома зазубрены.
F. vesiculosus (так называемые пузырчатые водоросли) - у высшей точки отлива. Имеются воздушные пузыри, которые обусловливают плавучесть. На рис. 3.18 можно видеть характерные внешние признаки F. vesiculosus, а на рис. 3.19 изображены основные особенности его внутреннего строения.

Рис. 3.18. Внешнее строение Fucus vesiculosus. Отмечены характерные признаки и, в частности, приспособления к окружающей среде. Плодущий конец
(рецептакул) представляет собой набухшую и покрытую мелкими вздутиями (скафидиями или концептакулами), сообщающимися с наружной средой только узкими отверстиями, часть таллома. У женских растений плодущие концы темно-зеленые, у мужских оранжевые. Воздушные пузыри
обычно парные и придают водоросли плавучесть. Придаточные ответвления
(иногда отламываются; это одна из форм вегетативного размножения). Верхушечная клетка
представляет собой точку роста, где происходит деление клеток. Ребро
- это жесткое образование, которое выполняет механические функции и, возможно, участвует в переносе некоторых веществ. Пластинка
плоская и упругая (кожистая); зеленовато-коричневого цвета из-за близкого к поверхности фотосинтезирующего слоя; покрыта слизью, предохраняющей от высыхания при отливе. Ребро вместе с пластинкой образуют таллом. Укореняющаяся часть таллома (в данном случае базальный диск
) бесцветна и очень прочно прикрепляет таллом к скалам и т. п. Размеры водоросли варьируют в пределах до 1 м или более. Слоевище
плоское и ремневидное; характер ветвления таков, что сопротивление волнам сводится к минимуму; воздушные пузыри поддерживают слоевище у поверхности, что способствует фотосинтезу. Черешок
- это в основном ребро; черешок гибкий и потому успешно противостоит волнам

В теле водоросли, или талломе, наблюдается некоторое разделение функций между разными тканями. Эта тенденция у Phaeophyta прослеживается лучше, чем у всех остальных групп водорослей. Приспособления водорослей к окружающей среде мы рассмотрим чуть позже.
Органы размножения
Половое размножение оогамное. F. vesiculosus и F. serratus - двудомные растения, т. е. у них есть и мужские, и женские особи. F. spiralis - гермафродит, у которого на одном растении в одних и тех же вместилищах - скафидиях, или концептакулах, - находятся и мужские, и женские репродуктивные органы. Репродуктивные органы развиваются внутри скафидиев на "плодущих" кончиках некоторых слоевищ. В каждом скафидии имеется узкое отверстие (пора), через которое впоследствии высвобождаются наружу репродуктивные органы. Их строение показано на рис. 3.19.
Взрослые растения диплоидные, а гаметы образуются в результате мейотического деления.
Приспособления к окружающей среде
Прежде чем мы рассмотрим приспособления Fucus к среде обитания, следует сказать несколько слов о самой среде, которая достаточно враждебна. Будучи растениями приливно-отливной зоны, разные водоросли в разной степени подвергаются воздействию воздушной среды во время отлива. Поэтому у них должны быть защитные приспособления от высыхания. К тому же и температура очень резко меняется, когда холодные морские волны вливаются в прогретые лужицы, оставшиеся после отлива. Растения должны быть приспособлены и еще к одному фактору, а именно к резким изменениям солености воды, будь то ее увеличение при испарении из небольших водоемов, образовавшихся после отлива, или ее уменьшение во время дождя. Для того чтобы противостоять таким факторам, как приливы, отливы, прибой и удары волн, нужна достаточная механическая прочность. Большие волны начинают перекатывать камни, и это может очень сильно повредить растения.
Морфологические приспособления (общее строение)
Слоевище водоросли прочно прикреплено к грунту укореняющейся частью таллома (ризоидами или базальным диском) (рис. 3.18). Оно настолько прочно связывается с грунтом (обычно это камни), что водоросль чрезвычайно трудно оторвать от него. Как правило, первым не выдерживает камень, а не укореняющаяся часть таллома.
Таллом водорослей не сплошной, а рассеченный; он дихотомически ветвится в одной плоскости, и это позволяет свести к минимуму сопротивление толще воды. К тому же он прочный и упругий, но не жесткий. Ребра слоевища крепкие и гибкие.
У плавучей водоросли F. vesiculosus имеются специальные воздушные пузыри, которые удерживают слоевище у поверхности воды, т. е. в условиях, способствующих максимальному улавливанию света для фотосинтеза.
Физиологические приспособления
Среди фотосинтетических пигментов преобладает бурый пигмент - фукоксантин . Это - одно из приспособлений к фотосинтезу под водой, так как фукоксантин сильно поглощает синий свет, который проникает в толщу воды гораздо дальше, чем более длинноволновые лучи, например красные.
Таллом выделяет много слизи, которая заполняет все внутренние полости водоросли и просачивается наружу. Слизь помогает лучше удержать воду и препятствует обезвоживанию.
Осмотическое давление в клетках намного выше, чем в морской воде, поэтому осмотических потерь воды не наблюдается.
Приспособления к половому размножению
Выход гамет синхронизирован с приливами. Во время отлива таллом обсыхает, и из скафидиев наружу выдавливаются репродуктивные органы, которые от высыхания предохраняются слизью. Во время прилива стенки репродуктивных органов растворяются, высвобождая гаметы. Мужские гаметы подвижны и обладают положительным хемотаксисом в отношении веществ, выделяемых женскими гаметами.
Развитие зиготы происходит сразу же после оплодотворения, что сводит к минимуму риск быть унесенной в океан.
3.2.6. Отдел Euglenophyta
Основные признаки Euglenophyta приведены в табл. 3.4. Для этого отдела характерны признаки как растений, так и животных, что сильно затрудняет классификацию относящихся сюда организмов. По этой причине их обычно включают в свои систематические схемы и ботаники, и зоологи. Эти проблемы мы обсудим позднее, после описания рода Euglena.
Euglena - самая обычная одноклеточная водоросль, живущая в пресноводных прудах, канавах и любых других водоемах, богатых растворенными органическими соединениями. Как и Chlamydomonas, она иногда размножается так интенсивно, что вода становится зеленой, потому что среди пигментов эвглены преобладает хлорофилл. Строение эвглены показано на рис. 3.20, где отмечены и некоторые ее особенности.

Рис. 3.20. Строение Euglena gracillis. Канал
- место, через которое поступает пища у незеленых видов; пелликула здесь отсутствует, что позволяет заглатывать мелкие частички. Глазок
(стигма) имеет красный цвет; участвует в реакции фототаксиса. Фоторецептор
обнаруживает источник света и заставляет организм плыть в направлении оптимальной освещенности (фототаксис); направление движения может меняться при затенении фоторецептора. Длинный жгутик
используется для локомоции; обычно направлен вперед; волнообразные движения проходят по жгутику от основания к кончику; жгутик тащит за собой клетку; во время движения вперед клетка вращается вокруг своей оси, оставляя за собой штопорообразный след. Пульсирующая вакуоль
окружена вспомогательными вакуолями; участвует в осморегуляции, выкачивая в резервуар избыток воды, поступившей в клетку в результате осмоса. Короткий жгутик
не участвует в локомоции. Парамилоновая гранула
образована полимером глюкозы, похожим на крахмал и являющимся запасным углеводом. Пелликула
располагается под плазматической мембраной
; гибкая. Хлоропласты
содержат фотосинтетические пигменты. В цитоплазме
находятся сократительные волокна, которые обусловливают перистальтические волны деформации клетки; такое движение называется эвгленоидным
У Euglena нет клеточной стенки. Снаружи клетка покрыта плазматической мембраной, сразу же под которой находится белковая пелликула . Пелликула довольно гибкая, и это позволяет клетке принимать разную форму. Пелликула полностью окружает цитоплазму, и ее можно рассматривать как своего рода наружный скелет . Она состоит из ряда утолщенных продольных полосок и микрофибрилл, переплетенных между собой. Когда внутри цитоплазмы сокращаются крошечные фибриллы, которые называются мионемами , полоски пелликулы начинают скользить относительно друг друга, в результате чего изменяется форма тела. Это явление называется эвгленоидным движением . Другой, более обычный для эвглены способ передвижения за счет вращения длинного жгутика изображен на рис. 3.20 (рассмотрите глазок, фоторецептор и длинный жгутик) и подробно описан в разд. 17.6.3.
Бесполое размножение происходит посредством продольного деления клетки надвое. Полового размножения не наблюдается.
Питание
Зеленые виды Euglena автотрофны и синтезируют все необходимые им вещества из двуокиси углерода, воды и минеральных солей. Вместе с тем они нуждаются в поступлении извне витаминов В1 и В12, которые они не могут синтезировать сами. В этом Euglena не отличается от животных, хотя такая потребность в витаминах характерна и для многих других водорослей.
У нескольких видов Euglena хлорофилла нет, и поэтому они не окрашены и не способны к фотосинтезу (т. е. гетеротрофны). Питаются они по типу сапрофитов, переваривание происходит вне клетки. Когда водоем загрязнен, они процветают, так как разлагающийся материал богат органическими соединениями. Другие бесцветные формы способны заглатывать мелкие частички пищи, для чего у них имеется своеобразная "глотка", где отсутствует пелликула. Затем эти частички перевариваются внутри клетки (голозойное питание, разд. 10.1.1). Пища загоняется в глотку за счет движения жгутиков. Эти виды во многом напоминают простейшее Реrапеmа (разд. 4.1.1).
Если зеленые клетки Euglena долго держать в темноте, то хлоропласты исчезают, и клетки становятся бесцветными. Если в среде достаточно органических веществ, то клетки могут долго жить как сапрофиты. Когда их переносят на свет, снова появляется хлорофилл.
Проблемы систематики Euglena
Как мы уже говорили и как это следует из табл. 3.1, для Euglena характерны признаки и растений, и животных. Один из таких животных признаков, который мы еще не рассматривали, - это наличие в глазке астаксантина - пигмента, свойственного животным.
Легкость, с которой некоторые эвглены могут переходить из зеленой формы в бесцветную и наоборот, свидетельствует о том, что постоянно бесцветные виды, по-видимому, произошли от зеленых. Если впоследствии у бесцветных форм возникли специальные приспособления для голозойного питания, подобные тем, которые имеются у Peranema, то вполне возможно, что предки простейших были похожи на растения. Не следует забывать, однако, что эволюция могла идти и в обратном направлении, ведь мы уже обсуждали в начале этой главы возможность того, что предки растений могли быть похожи на животных (т. е. на гетеротрофных эукариот).
Решая вопрос о том, в царство растений или в царство животных помещать Euglena, необходимо помнить, что некоторые признаки животных имеются и у хламидомонады, и тем не менее ее обычно относят к растениям. Основные затруднения систематиков связаны со способом питания. Судя по всему, эвглену все же следует относить к растениям, так как наличие хлоропластов считается уникальной особенностью, присущей только царству растений. Все это, однако, лишний раз напоминает нам, сколь трудно навязать природе искусственную систематику, придуманную людьми.
3.3. Составьте таблицу растительных и животных признаков Euglena. Воспользуйтесь для этого табл. 3.1, рис. 3.20 и сведениями, изложенными выше.
3.2.7. Направления эволюции водорослей
Даже тех нескольких примеров, которые мы рассмотрели в предыдущих разделах, вполне достаточно, чтобы понять, что существует множество типов водорослей, включая и такие одноклеточные формы, как Chlamydomonas, и такие сравнительно крупные организмы, как Fucus, у которых тело дифференцировано и наблюдается определенное разделение функций между отдельными тканями. У некоторых крупных бурых водорослей имеются даже проводящие ткани, хотя настоящей проводящей ткани - ксилемы и флоэмы - у них нет.
У водорослей четко прослеживается тенденция к усложнению процесса полового размножения от простой изогамии и анизогамии к оогамии. Однако следует с большой долей осторожности использовать ту или иную тенденцию для объяснения эволюционных взаимосвязей между отдельными группами водорослей. Такие взаимосвязи до сих пор окончательно не выяснены, а группа Chlorophyta (зеленые водоросли), от которой, как полагают, произошли наземные растения, отличается очень большим разнообразием: в ней есть и простые одноклеточные формы, и гораздо более сложные, а половое размножение также варьирует от изогамии до оогамии.
3.2.8. Значение водорослей
Роль водорослей в биосфере
По современным оценкам, на долю океана приходится по меньшей мере половина мировой первичной продукции, выражающейся в количестве фиксированного углерода. Эту первичную продукцию образуют водоросли - единственные растения, которые населяют океан. Учитывая ту огромную площадь, которую занимает океан, следует ожидать, что его продуктивность должна быть еще больше, но нельзя забывать, что фотосинтез возможен только в поверхностных слоях, куда проникает свет и где лимитирующим фактором является доступность биогенных элементов, особенно азота и фосфора.
Водоросли - очень важные первичные продуценты (гл. 12), с которых начинается большинство пищевых цепей, в том числе практически все морские и многие пресноводные цепи. Эти цепи через зоопланктон * , ракообразных и т. п. доходят до рыб. Многие микроскопические водоросли - одноклеточные, и именно они являются главным компонентом фитопланктона * .
* (Планктон это мельчайшие растения (фитопланктон) и животные (зоопланктон), которые свободно плавают в поверхностных слоях океанов и озер. Планктон имеет очень большое хозяйственное и экологическое значение. )
Фиксация углерода - это только одно из следствий фотосинтеза (разд. 9.2). Кроме того, благодаря фотосинтезу поддерживается уровень кислорода в атмосфере, при этом по меньшей мере половину всего кислорода выделяют водоросли, и их вклад в этот процесс намного больше, чем вклад наземных лесов.
Альгиновая кислота, агар и каррагенан
Из водорослей получают многие полезные продукты, например альгиновую кислоту, агар и каррагенан. Альгиновая кислота и ее производные (альгинаты) - это полисахариды, которые экстрагируют из срединной пластинки и клеточных стенок таких бурых водорослей, как Laminaria, Ascophyllum и Macrocystis. Водоросли добывают в большом количестве в прибрежных мелких водах; Macrocystis, например, собирают на побережье Калифорнии. Очищенные альгинаты не токсичны и легко образуют гели. Их широко применяют в качестве отвердителей и желеобразующих веществ для получения промышленных товаров (например, в косметике - для изготовления кремов для рук); в качестве эмульгаторов - для приготовления мороженого; в качестве желеобразующих веществ - в кондитерской промышленности; при изготовлении лаков, красок и лекарств; для получения глазурованной керамической посуды.
Агар - полисахарид, который получают из красных водорослей. Он образует такие же гели, как и альгинаты, но, возможно, более известен, так как является очень удобной средой для выращивания бактерий и грибов. С этой целью готовят разбавленный раствор агара, затем добавляют в него различные питательные вещества, стерилизуют и дают застыть, получая желеобразную массу. Кроме того, агар используют для тех же самых целей, что и альгинаты.
Каррагенан (карраген) - это еще один полисахарид клеточной стенки, который получают главным образом из красной водоросли Chondrus crispus. По своей химической структуре он очень похож на агар и применяется для тех же самых целей.
Диатомит (кизельгур)
Водоросли, относящиеся к отделу Bacillariophyta, в основном одноклеточные; их называют диатомовыми . Для этих водорослей характерно особое строение клеточной стенки, в которой содержится кремний. После гибели клеток остатки диатомей падают на дно морей и озер, и постепенно там накапливаются большие отложения. Образующаяся таким образом "диатомовая земля" содержит очень много (до 90%) кремния. После соответствующей очистки эту "землю" можно использовать как превосходный фильтрующий материал (например, при получении сахара или для осветления пива), как наполнитель при изготовлении красок или бумаги и как изоляционный материал, способный противостоять резким перепадам температуры.
Удобрение
На фермах, расположенных вблизи побережья, крупные водоросли (красные и бурые) по традиции используют как удобрения, хотя и в незначительных масштабах. Водоросли богаты калием, но в них гораздо меньше азота и фосфора, чем в простом навозе. Поэтому их удобряющее действие не очень велико. Более значимую роль играют свободноживущие сине-зеленые водоросли, которые являются очень важными азотфиксаторами и довольно распространены в почве (разд. 9.11.1).
Пищевые продукты
Некоторые водоросли подают прямо к столу, особенно на Дальнем Востоке. Красную водоросль Porphyra, считающуюся деликатесом, и большую бурую водоросль Laminaria обычно едят в сыром виде или же готовят из них разные блюда. В Южном Уэльсе Porphyra кладут в одно из традиционных блюд, для приготовления которого отваренные водоросли смешивают с овсянкой и потом все это тушат в масле. В поисках новых источников пищи много внимания было уделено промышленному культивированию водорослей. Однако для получения новых пищевых продуктов годятся очень немногие водоросли, и до сего времени сколько-нибудь значимые успехи в этой области были достигнуты при культивировании бактерий и грибов. Из сине-зеленых водорослей многообещающей считается Spirulina.
Очистка сточных вод
Водоросли вносят определенный вклад в работу микроорганизмов по очистке сточных вод, так как в сточных водах содержатся питательные вещества не только для бактерий, грибов и простейших, но и для микроскопических зеленых водорослей. Они особенно полезны в открытых "окислительных прудах", которые достаточно широко используются в тропических и субтропических странах. Открытые пруды глубиной от 1 до 1,5 м заливают неочищенными стоками. В процессе фотосинтеза водоросли выделяют кислород и обеспечивают таким образом жизнедеятельность других аэробных микроорганизмов, растущих в сточных водах. Время от времени водоросли собирают и перерабатывают на корм скоту.
Научные исследования
Одноклеточным водорослям присущи все характерные признаки типичных растений, поэтому они являются идеальным материалом для научных исследований, так как, во-первых, их можно выращивать в большом количестве в строго определенных условиях и, во-вторых, для этого не требуется очень много места. Примером таких водорослей может служить Chlorella, которой по праву принадлежит почетное место в исследованиях фотосинтеза (разд. 9.4.3). Водоросли используются и при изучении поглощения ионов. Они принесли большую пользу и в новаторских исследованиях строения клеточной стенки и жгутиков.
Вред, наносимый водорослями
В определенных условиях водоросли "цветут", т. е. в огромных количествах скапливаются в воде. "Цветение" наблюдается при достаточно теплой погоде, когда в воде много питательных веществ. Такая ситуация очень часто искусственно создается человеком, когда в воду сбрасывают промышленные стоки или же когда в реки и озера попадают удобрения с полей. В результате начинается взрывоподобное размножение первичных продуцентов (водорослей), и они в нарушение всех законов природы начинают отмирать раньше, чем их успеют съесть. При последующем разложении остатков происходит столь же интенсивное размножение аэробных бактерий и вода полностью лишается кислорода. Все это происходит очень быстро, и из-за нехватки кислорода начинают гибнуть рыбы и другие животные и растения. Увеличение концентрации питательных веществ в воде, которое запускает весь этот процесс, называется эвтрофизацией водоема, и если оно происходит быстро, то можно считать, что это еще одна из форм загрязнения окружающей среды.
Токсины, образующиеся при "цветении" воды, в особенности при размножении сине-зеленых водорослей, увеличивают гибель животных. Подобные взрывы численности водорослей представляют собой серьезную проблему для рыбоводческих хозяйств, особенно там, где интенсивный вывоз удобрений на поля еще в большей степени усиливает эвтрофизацию. Сходные осложнения возникают и при "цветении" воды в океане. Кроме того, токсины, накапливаясь в теле моллюсков и ракообразных, питающихся водорослями, и затем попадая в организм человека, вызывают у него различные отравления и паралич.
С водорослями связаны и многие сложности при хранении питьевой воды в запасных резервуарах, когда она загрязняется продуктами жизнедеятельности водорослей или когда водоросли начинают расти на песчаных фильтрах, полностью забивая их.
3.4. Те трудности, о которых мы только что говорили, чаще возникают в водохранилищах, расположенных в низинах. Объясните, почему это так и должно быть.
3.5. В отличие от многих грибов и бактерий водоросли не вызывают никаких заболеваний. С чем это связано?
Водоросли - большая группа древнейших растений. Строение их тела и размеры характеризуются огромным разнообразием. Существуют микроскопических размеров одноклеточные, многоклеточные и колониальные формы (в 1-2мкм) и крупные, с различным строением слоевища, достигающие 30-45м. Рассмотрим общую характеристику водорослей.
Общим свойством всех водорослей является наличие хлорофилла. Кроме хлорофилла водоросли могут содержать и другие пигменты (фикоциан, фикоэритрин, каротин, ксантофилл, фикоксантин). Эти пигменты придают водорослям красную, бурую, желто-зеленую окраску, маскируя основную зеленую.
Общая характеристика
Питание . Наличие пигментов в клетках водорослей обеспечивает аутотрофный тип питания. Однако многие водоросли обладают способностью в определенных условиях переключаться на гетеротрофное питание (эвгленовые - в темноте) либо сочетают его с фотосинтезом.
Классификация водорослей . Количество видов водорослей превышает 40тыс. Однако классификация их не завершена, так как не все формы достаточно хорошо изучены. У нас в стране принято деление водорослей на 10 отделов: синезеленые, пирофитовые, золотистые, диатомовые, желтозеленые, бурые, красные, эвгленовые, зеленые, харовые. Наибольшим числом видов представлены зеленые (13-20 тыс.) и диатомовые (10 тыс.).
Деление водорослей на отделы совпадает обычно с их окраской, связанной, как правило, с особенностями строения клеток и слоевища.
Строение клеток водоросли . Водоросли - единственная группа организмов, среди которых встречаются прокариоты (синезеленые) и эукариоты (все остальные). В ядрах эукариотических водорослей выявлены все структуры, свойственные ядрам других эукариот: оболочки, ядерный сок, ядрышки, хромосомы.

Строение, состав и свойства остальных клеточных компонентов у водорослей характеризуется большим разнообразием. В процессе эволюции естественный отбор сохранил наиболее перспективные формы, в том числе такой тип клеточной организации, который позволил растениям перейти к наземному образу жизни.
Размножение водорослей бывает вегетативным, бесполым (с помощью спор) и половым. У одного и того же вида в зависимости от условий и времени года способы размножения различны. При этом наблюдается смена ядерных фаз - гаплоидной и диплоидной.
Условия жизни и места обитания водорослей . Благоприятными условиями существования водорослей являются: наличие света, источников углерода и минеральных солей, а основной средой обитания для них служит вода. Большое влияние на жизнь водорослей оказывают температура, соленость воды и др.
Большинство водорослей - обитатели пресных и морских водоемов. Они могут населять толщу воды, свободно плавать в ней, образуя фитопланктон, либо поселяются на дне, прикрепляясь к разнообразным предметам, живым и мертвым организмам, образуя фитобентос. Заселяют водоросли и горячие источники, а также водоемы с повышенной соленостью.