Quali cellule contengono lisosomi? Struttura e funzioni dei lisosomi
In una gabbia. Inoltre, presteremo attenzione allo scopo di questo organello e alla sua struttura.
Come è già chiaro, il lisosoma è parte integrante di ogni cellula. E tutto ciò che vediamo, ciò che tocchiamo e noi stessi siamo un costruttore costituito da tante minuscole particelle. La cellula è l'unità strutturale elementare di tutti gli organismi che vivono sul pianeta Terra. Ha una serie di qualità che le permettono di esistere in modo indipendente:
- proprio metabolismo;
- capacità di riprodursi;
- riproduzione (autoriproduzione);
- sviluppo.
Bene, ora proponiamo di passare all'organello che ci interessa, considerarne la struttura ed evidenziare le funzioni dei lisosomi nella cellula.
Lisosoma
Ora analizzeremo questo organello in modo più dettagliato e ti forniremo una classificazione. Prima di elencare e rivedere le funzioni dei lisosomi nella cellula, è necessario menzionare una breve storia della scoperta. Questi piccoli granuli furono scoperti per la prima volta dallo scienziato de Duve in una cellula del fegato. Questo evento si è verificato negli anni '50 del XX secolo.
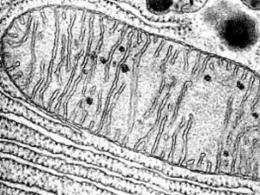
Un lisosoma è una cavità piena di vari enzimi idrolitici (ne esistono più di 80 tipi). È circondato da una membrana, è importante chiarire che è unica. L'aspetto di questi organelli non è lo stesso; nella maggior parte dei casi è di forma rotonda, con un diametro non superiore a 0,8 micron.
La membrana del lisosoma non ha lo stesso spessore, la sua permeabilità cambia sotto l'influenza di determinate condizioni. Quindi, i labilizzatori (cioè aumentando la permeabilità) sono:
- tiroxina;
- progesterone;
- vitamina A;
- raggi ultravioletti;
- Radiazione a raggi X;
- ossigeno, ecc.
L'effetto opposto è esercitato da:
- prednisolone;
- cortisone, ecc.
Cellule diverse non hanno lo stesso numero di lisosomi; la maggior parte di essi si trova in cellule che hanno la funzione di fagocitosi. Gli esempi includono macrofagi o leucociti. Ciò include anche quelli che sono capaci di assorbimento, secrezione ed escrezione. Sono:
- cellule epiteliali;
- intestini;
- rene;
- ghiandola prostatica, ecc.
Ora brevemente sulla classificazione dei lisosomi. Ne esistono di due tipi: primari e secondari. Quelli primari sono detti cumulativi. Tra quelli secondari possiamo evidenziare:
- fagolisosomi;
- citolisosomi;
- corpi residui.

Funzioni
Proponiamo ora di evidenziare le poche funzioni dei lisosomi nella cellula. Quindi, questo include:
- digestione cellulare;
- autofagia;
- autolisi;
- dissoluzione delle strutture esterne.
Spiegheremo ora brevemente il significato di questi termini. Puoi leggere in dettaglio sulla digestione cellulare e sull'autofagia un po 'più tardi. Ora parliamo della funzione svolta dai lisosomi durante la morte cellulare.
Questo processo è comunemente chiamato autolisi. La membrana del lisosoma può essere rotta, il che porta al rilascio degli enzimi in essa contenuti. Di norma, cessano di svolgere la loro funzione principale, poiché vengono semplicemente disattivati nel citoplasma della cellula.
La distruzione di una cellula non è un problema, ma cosa succede se tutti i lisosomi interrompono la loro struttura? Quindi si verifica la morte della cellula stessa. Un esempio lampante di autolisi è la morte della coda nei girini di rana.

Digestione
Abbiamo accennato in precedenza che i lisosomi eseguono la digestione. Ti invitiamo a considerare questo processo in modo più dettagliato. Come già accennato, i lisosomi si dividono in due gruppi, tra cui secondario è anche il vacuolo digestivo. È lei che svolge la funzione di digestione nella cellula. È formato dalla fusione di un fagosoma e di un lisosoma primario.
Il vacuolo digestivo ha dimensioni maggiori, arrivando fino a 1,2 micron. Contiene un numero molto elevato di inclusioni. È qui che avviene l'elaborazione delle sostanze che entrano nella cellula. Accade spesso che vengano digeriti mediante idrolisi in particelle a basso peso molecolare. Quest'ultimo può facilmente passare attraverso la membrana lisosomiale. Inoltre, la cellula ne ha bisogno per formare nuovi organelli.
Autofagia

Quale altra funzione svolgono i lisosomi in una cellula? Abbiamo già detto che tra i loro scopi c'è qualcosa come l'autofagia. Questo processo è caratterizzato dalla cattura di componenti nella cellula e dalla distruzione da parte dei lisosomi. Esistono 3 tipi di autofagia:
- micro;
- macro;
- accompagnatore
Nel primo caso, il lisosoma cattura i detriti e li digerisce per produrre energia o materiale da costruzione. Questo processo può verificarsi durante il digiuno. Durante la macroautofagia, l’autofagosoma e il lisosoma si uniscono, dando luogo alla formazione di un autofagolisosoma. In quest'ultimo vengono digeriti i resti dei futofagosomi. Il terzo tipo può essere osservato esclusivamente nei mammiferi durante i periodi di stress. Con questo tipo di autofagia avviene il trasporto mirato di proteine nei lisosomi.
Diversi tipi di lisosomi possono essere considerati compartimenti cellulari separati.
I lisosomi furono scoperti nel 1955 dal biochimico belga Christian de Duve. I lisosomi sono presenti in tutte le cellule dei mammiferi, ad eccezione dei globuli rossi. Nelle piante, i vacuoli sono simili ai lisosomi nel metodo di formazione e in parte nella funzione. I lisosomi sono presenti anche nella maggior parte dei protisti (sia con alimentazione fagotrofica che osmotrofica) e nei funghi. Pertanto, la presenza di lisosomi è caratteristica delle cellule di tutti gli eucarioti. I procarioti non hanno lisosomi perché mancano di fagocitosi e non hanno digestione intracellulare.
Numerose malattie ereditarie nell’uomo, chiamate malattie da accumulo lisosomiale, sono associate alla disfunzione dei lisosomi.
Storia della scoperta
Nel 1974 C. de Duve ricevette il Premio Nobel per la Medicina per il suo contributo alla scoperta dell'organizzazione strutturale e funzionale della cellula.
Segni di lisosomi
I lisosomi sono eterogenei per forma, dimensione, caratteristiche ultrastrutturali e citochimiche. Nelle cellule animali, i lisosomi hanno in genere dimensioni inferiori a 1 µm, sebbene in alcuni tipi di cellule, come i macrofagi, i lisosomi possono essere più grandi di diversi micron. I lisosomi, di regola, hanno una forma sferica, ovale, a volte tubolare. Il numero di lisosomi varia da uno (un grande vacuolo in molte cellule vegetali e fungine) a diverse centinaia o migliaia (nelle cellule animali). I lisosomi negli animali di solito non costituiscono più del 5% del volume intracellulare.
Una delle caratteristiche dei lisosomi è la presenza in essi di numerosi enzimi (idrolasi acide) in grado di scomporre proteine, carboidrati, lipidi e acidi nucleici. Gli enzimi lisosomiali includono catepsine (proteasi tissutali), ribonucleasi acida, fosfolipasi, ecc. Inoltre, i lisosomi contengono enzimi in grado di rimuovere gruppi solfato (solfatasi) o fosfato (fosfatasi acida) da molecole organiche. In totale, la cavità del lisosoma contiene circa 60 enzimi idrolitici acidi solubili.
I lisosomi sono caratterizzati da una reazione acida dell'ambiente interno, che garantisce il funzionamento ottimale delle idrolasi lisosomiali. Di solito nei lisosomi è circa 4,5-5, cioè la concentrazione di protoni in essi è due ordini di grandezza superiore rispetto al citoplasma. Ciò è assicurato dal trasporto attivo dei protoni, effettuato dalla proteina della pompa protonica ATPasi incorporata nelle membrane dei lisosomi. Oltre alla pompa protonica, nella membrana del lisosoma sono integrate proteine trasportatrici per il trasporto dei prodotti dell'idrolisi delle macromolecole nel citoplasma: aminoacidi, zuccheri, nucleotidi, lipidi.
L'elevata attività della fosfatasi acida è stata precedentemente utilizzata come uno dei marcatori dei lisosomi. Attualmente, la presenza di glicoproteine di membrana specifiche - LAMP1 e LAMP2 è considerata un marcatore più affidabile. Sono presenti sulla membrana dei lisosomi e degli endosomi tardivi, ma sono assenti sulle membrane degli altri compartimenti del vacuoma.
Formazione dei lisosomi e loro tipi
I lisosomi sono formati da vescicole (vescicole) che si separano dall'apparato del Golgi e vescicole (endosomi) in cui entrano sostanze durante l'endocitosi. Le membrane del reticolo endoplasmatico partecipano alla formazione degli autolisosomi (autofagosomi). Tutte le proteine lisosomiali sono sintetizzate sui ribosomi sessili sul lato esterno delle membrane del reticolo endoplasmatico e poi passano attraverso la sua cavità e attraverso l'apparato di Golgi.
Non esiste una classificazione e nomenclatura generalmente accettata per i diversi stadi di maturazione e tipi di lisosomi. Esistono lisosomi primari e secondari. I primi si formano nella regione dell'apparato del Golgi, contengono enzimi allo stato inattivo, mentre i secondi contengono enzimi attivi. Tipicamente, gli enzimi lisosomiali vengono attivati quando il pH diminuisce. Tra i lisosomi si possono distinguere anche gli eterolisosomi (che digeriscono il materiale che entra nella cellula dall'esterno - attraverso la fago o la pinocitosi) e gli autolisosomi (che distruggono le proteine o gli organelli propri della cellula). La classificazione più utilizzata dei lisosomi e dei compartimenti associati è:
- L'endosoma precoce: vi entrano vescicole endocitiche (pinocitotiche). Dall'endosoma iniziale, i recettori che hanno ceduto il loro carico (a causa del pH basso) ritornano alla membrana esterna.
- Endosoma tardivo: vescicole con materiale assorbito durante la pinocitosi e vescicole dell'apparato di Golgi con idrolasi entrano dall'endosoma precoce. I recettori del mannosio 6-fosfato ritornano dall'endosoma tardivo all'apparato di Golgi.
- Lisosoma: le vescicole con una miscela di idrolasi e materiale digeribile entrano dall'endosoma tardivo.
- Fagosoma: particelle più grandi (batteri, ecc.) entrano al suo interno e vengono assorbite dalla fagocitosi. I fagosomi di solito si fondono con un lisosoma.
- Un autofagosoma è una regione del citoplasma circondata da due membrane, che solitamente comprende alcuni organelli e che si forma durante la macroautofagia. Si fonde con il lisosoma.
- Corpi multivescicolari - solitamente circondati da un'unica membrana, contengono al loro interno vescicole più piccole circondate da un'unica membrana. Formato da un processo che ricorda la microautofagia (vedi sotto), ma contenente materiale ottenuto dall'esterno. Nelle piccole vescicole, i recettori della membrana esterna (ad esempio, i recettori del fattore di crescita epidermico) solitamente rimangono e vengono poi degradati. Lo stadio di formazione corrisponde ai primi endosomi. È stata descritta la formazione di corpi multivescicolari circondati da due membrane per gemmazione dall'involucro nucleare.
- I corpi residui (telolisosomi) sono vescicole contenenti materiale non digerito (in particolare lipofuscina). Nelle cellule normali si fondono con la membrana esterna e lasciano la cellula per esocitosi. Si accumulano con l'invecchiamento o con la patologia.
Funzioni dei lisosomi
Le funzioni dei lisosomi sono:
- digestione di sostanze o particelle catturate dalla cellula durante l'endocitosi (batteri, altre cellule)
- autofagia - la distruzione di strutture non necessarie per la cellula, ad esempio durante la sostituzione di vecchi organelli con nuovi, o la digestione di proteine e altre sostanze prodotte all'interno della cellula stessa
- l'autolisi è l'autodigestione di una cellula, che porta alla sua morte (a volte questo processo non è patologico, ma accompagna lo sviluppo dell'organismo o la differenziazione di alcune cellule specializzate). Esempio: Quando un girino si trasforma in una rana, i lisosomi situati nelle cellule della coda lo digeriscono: la coda scompare e le sostanze formate durante questo processo vengono assorbite e utilizzate da altre cellule del corpo.
- dissoluzione delle strutture esterne (vedi, ad esempio, gli osteoclasti)
Digestione intracellulare e partecipazione al metabolismo
In molti protisti e animali con digestione intracellulare, i lisosomi sono coinvolti nella digestione del cibo catturato mediante endocitosi. In questo caso i lisosomi si fondono con i vacuoli digestivi. Nei protisti, i resti di cibo non digerito vengono solitamente rimossi dalla cellula attraverso la fusione del vacuolo digestivo con la membrana esterna.
Molte cellule animali in cui predomina la digestione della cavità (ad esempio i cordati) ricevono nutrienti dal fluido intercellulare o dal plasma sanguigno mediante la pinocitosi. Queste sostanze sono coinvolte anche nel metabolismo cellulare dopo essere state digerite nei lisosomi. Un esempio ben studiato di tale partecipazione dei lisosomi al metabolismo è la produzione di colesterolo da parte delle cellule. Il colesterolo, trasportato dal sangue sotto forma di LDL, entra nelle vescicole pinocitotiche dopo che l'LDL si lega ai recettori LDL sulla membrana. I recettori ritornano alla membrana dall'endosoma iniziale e le LDL entrano nei lisosomi. Successivamente, l'LDL viene digerito e il colesterolo rilasciato entra nel citoplasma attraverso la membrana del lisosoma.
Indirettamente, i lisosomi partecipano al metabolismo, fornendo la desensibilizzazione delle cellule agli effetti degli ormoni. Con l'azione prolungata di un ormone su una cellula, alcuni dei recettori che legano l'ormone entrano negli endosomi e poi si degradano all'interno dei lisosomi. Una diminuzione del numero di recettori riduce la sensibilità della cellula all'ormone.
Autofagia
Esistono solitamente due tipi di autofagia: microautofagia e macroautofagia. Durante la microautofagia, come nella formazione dei corpi multivescicolari, si formano invaginazioni della membrana dell'endosoma o del lisosoma, che vengono poi separate sotto forma di vescicole interne, in cui entrano solo le sostanze sintetizzate nella cellula stessa. In questo modo, la cellula può digerire le proteine quando manca energia o materiale da costruzione (ad esempio durante la fame). Ma i processi di microautofagia si verificano anche in condizioni normali e generalmente non sono selettivi. A volte gli organelli vengono anche digeriti durante la microautofagia; Pertanto, nel lievito sono state descritte la microautofagia dei perossisomi e la microautofagia parziale dei nuclei, in cui la cellula rimane vitale.
Nella macroautofagia, una regione del citoplasma (spesso contenente qualche tipo di organello) è circondata da un compartimento di membrana simile al serbatoio del reticolo endoplasmatico. Di conseguenza, quest'area è separata dal resto del citoplasma da due membrane. Questo autofagosoma si fonde quindi con il lisosoma e il suo contenuto viene digerito. Apparentemente, anche la macroautofagia non è selettiva, sebbene venga spesso sottolineato che con il suo aiuto la cellula può liberarsi di organelli "obsoleti" (mitocondri, ribosomi, ecc.).
Il terzo tipo di autofagia è dipendente dall'accompagnatore. Con questo metodo, il trasporto diretto delle proteine parzialmente denaturate dal citoplasma avviene attraverso la membrana del lisosoma nella sua cavità.
Autolisi
Gli enzimi lisosomiali vengono spesso rilasciati quando la membrana lisosomiale viene distrutta. Di solito vengono inattivati nell'ambiente neutro del citoplasma. Tuttavia, con la distruzione simultanea di tutti i lisosomi della cellula, può verificarsi la sua autodistruzione: l'autolisi. Esistono autolisi patologiche e ordinarie. Una variante comune dell'autolisi patologica è l'autolisi tissutale post-mortem.
Normalmente, i processi di autolisi accompagnano molti fenomeni associati allo sviluppo del corpo e alla differenziazione cellulare. Pertanto, l'autolisi cellulare è descritta come un meccanismo di distruzione dei tessuti nelle larve di insetti durante la metamorfosi completa, nonché durante il riassorbimento della coda di un girino. È vero che queste descrizioni si riferiscono ad un periodo in cui le differenze tra apoptosi e necrosi non erano ancora state stabilite, e in ogni caso bisogna verificare se l'apoptosi, non associata all'autolisi, è effettivamente alla base della degradazione di un organo o tessuto.
Nelle piante l'autolisi è accompagnata dalla differenziazione delle cellule che funzionano dopo la morte (ad esempio tracheidi o segmenti vascolari). L'autolisi parziale si verifica anche durante la maturazione delle cellule del floema - segmenti di tubi del setaccio.
Significato clinico
A volte, a causa del funzionamento improprio dei lisosomi, si sviluppano malattie da accumulo, in cui gli enzimi non funzionano o funzionano male a causa di mutazioni. Esempi di malattie da accumulo lisosomiale includono la malattia di Gaucher, la malattia di Pompe e la malattia di Tay-Sachs. In totale, sono note più di 50 malattie ereditarie associate alla ridotta funzionalità dei lisosomi.
Il danno ai lisosomi delle cellule necrotiche, compresi i granulociti, dà origine al processo infiammatorio.
Guarda anche
Appunti
- , Con. 1196.
- Settembre C. et al. Segnali dal lisosoma: un centro di controllo per la clearance cellulare e il metabolismo energetico. (Inglese) // Naz. Rev. Mol. Biolo cellulare. - 2013. -Vol. 14. - P. 283-296. -DOI:10.1038/nrm3565.
- Brighouse A., Dacks J.B., Field M.C. Evoluzione delle proteine Rab e storia del sistema endomembrana eucariotico (inglese) // Scienze della vita cellulare e molecolare. - 2010. -Vol. 67, n. 20 . - P. 3449-3465. -DOI:10.1007/s00018-010-0436-1. Archiviata dall'originale il 6 gennaio 2015.
- De Duve C. Il lisosoma compie cinquant'anni (inglese) // Natura biologia cellulare. - 2005. -Vol. 7, n. 9 . - P. 847-849.
- Lüllmann-Rauch R. Storia e morfologia del lisosoma// Lisosomi / P. Saftig. - Springer Stati Uniti, 2005. - P. 1-16. - ISBN 978-0-387-25562-0.
- Platt F.M., Boland B., van der Spoel A.C. Disturbi da accumulo lisosomiale: l'impatto cellulare della disfunzione lisosomiale // Il giornale della biologia cellulare. - 2012. -Vol. 199, n. 5 . - P.723-734. -DOI:10.1083/jcb.201208152.
- De Duve C. et al. Studi sul frazionamento dei tessuti. 6. Modelli di distribuzione intracellulare degli enzimi nel tessuto del fegato di ratto (inglese) // Biochemical Journal. - 1955. -Vol. 60, n. 4 . - P. 604–617.
Che si trovano nella maggioranza e svolgono la funzione di enzimi digestivi.
Cosa sono i lisosomi?
I lisosomi sono sacche di membrana sferica contenenti enzimi idrologici acidi in grado di digerire le macromolecole cellulari. La membrana lisosomiale aiuta a mantenere l'ambiente acido all'interno dell'organello e separa gli enzimi digestivi dal resto.
Gli enzimi lisosomiali sono prodotti da proteine del reticolo endoplasmatico e sono racchiusi in vescicole.
Enzimi lisosomiali
I lisosomi contengono vari enzimi idrolitici (circa 50 tipi diversi) che sono in grado di digerire acidi nucleici, polisaccaridi, lipidi e proteine. L'interno del lisosoma è costantemente mantenuto acido, poiché gli enzimi funzionano meglio in un ambiente acido. Se l'integrità del lisosoma è compromessa, gli enzimi non causeranno danni significativi al citosol neutro della cellula.
Formazione scolastica
I lisosomi si formano dalla fusione delle vescicole del complesso del Golgi con gli endosomi. Gli endosomi sono vescicole che si formano quando un sito viene bloccato e interiorizzato dalla cellula. In questo processo, il materiale extracellulare viene assorbito nella cellula. Quando gli endosomi maturano, diventano noti come endosomi tardivi.
Gli endosomi tardivi si fondono con le vescicole di trasporto del Golgi, che contengono idrolasi acide. Dopo la fusione, questi endosomi diventano lisosomi.
Funzioni dei lisosomi
I lisosomi agiscono come spazzini della cellula. Partecipano al riciclaggio del materiale cellulare organico e alla digestione intracellulare delle macromolecole.
Alcune cellule, come i globuli bianchi, hanno molti più lisosomi di altre. Queste cellule distruggono cellule, cellule morte, cellule tumorali e sostanze estranee attraverso la digestione cellulare. inghiottiscono una sostanza mediante fagocitosi e la racchiudono in una vescicola chiamata fago.
I lisosomi nel macrofago si fondono con il fagosoma, rilasciando i loro enzimi e formando il cosiddetto fagolisosoma. Il materiale interiorizzato viene digerito nel fagolisosoma. I lisosomi sono necessari anche per la degradazione dei componenti cellulari interni come gli organelli. In molti organismi, i lisosomi sono coinvolti nella morte cellulare programmata.
Difetti del lisosoma
Negli esseri umani, i lisosomi possono essere influenzati da varie condizioni ereditarie. Queste mutazioni genetiche causano malattie come la sindrome di Pompe, la sindrome di Hurler e la malattia di Tay-Sachs. Alle persone con questi disturbi mancano uno o più enzimi idrolitici lisosomiali. Ciò porta all'interruzione del normale metabolismo delle macromolecole nel corpo.
Lisosomi sono parte integrante della composizione cellulare. Sono un tipo di vescicola. Questi aiutanti cellulari, essendo parte del vacuoma, sono ricoperti da una membrana e pieni di enzimi idrolitici. L'importanza dell'esistenza dei lisosomi all'interno della cellula è assicurata dalla funzione secretoria, necessaria nel processo di fagocitosi e autofagocitosi.
I lisosomi esistono nelle cellule animali e vegetali e persino nelle cellule fungine. La struttura e le funzioni dei lisosomi possono differire. Possono avere diversi:
- dimensioni;
- forma;
- caratteristiche strutturali;
- caratteristiche della composizione chimica.
In una versione generalizzata lisosomi- si tratta di vescicole di membrana formate da vescicole ed endosomi, con ambiente acido (PH da 1,5 a 4-4,5).
Una caratteristica distintiva della struttura e delle funzioni delle vescicole di membrana, oltre alla reazione acida dell'ambiente interno, da altri organelli intracellulari è la presenza di enzimi proteici. Gli enzimi sono coinvolti nella scomposizione dei polimeri presenti nel corpo, vale a dire proteine, carboidrati, grassi e acidi nucleici. Le loro dimensioni variano da 0,3 a 0,5 micron, a seconda del loro habitat.
Funzioni dei lisosomi
Un insieme così ampio di enzimi all'interno di una vescicola microscopica è dovuto a una serie di importanti funzioni dei lisosomi eseguite nel corpo.
1. Negli organismi viventi che hanno una digestione intracellulare, prendono parte alla digestione dei nutrienti che entrano nella cellula durante il processo di endocitosi. Durante l'endocitosi, la cellula cattura non solo i nutrienti, ma anche i batteri che entrano nel corpo. Gli enzimi lisosomiali digeriscono le particelle catturate, fondendosi con i vacuoli.

I loro enzimi possono avere effetti benefici sulla cellula e sull'organismo nel suo insieme.
2. I lisosomi digeriscono non solo le sostanze necessarie, ma distruggono anche gli elementi dannosi che non sono necessari per la cellula (partecipano al rinnovamento cellulare). Gli enzimi scompongono anche i composti proteici prodotti dalla cellula stessa. Questo processo è chiamato autofagia dai biologi.
3. Durante l'autolisi, le cellule si autodistruggono, il che può avere un effetto dannoso sul corpo. Questo processo avviene durante lo sviluppo dell'organismo e la differenziazione delle cellule responsabili di processi specializzati.

4. Le funzioni dei lisosomi includono anche la loro partecipazione al metabolismo. Le sostanze digerite dalle bolle di membrana entrano nel fluido intercellulare o nel plasma sanguigno e sono coinvolte nel metabolismo.
Organidi- componenti permanenti, necessariamente presenti, della cellula che svolgono funzioni specifiche.
Reticolo endoplasmatico
Reticolo endoplasmatico (RE), O reticolo endoplasmatico (RE), è un organello a membrana singola. Si tratta di un sistema di membrane che formano “cisterne” e canali, collegati tra loro e delimitanti un unico spazio interno: le cavità dell'EPS. Le membrane sono collegate da un lato alla membrana citoplasmatica e dall'altro alla membrana nucleare esterna. Esistono due tipi di EPS: 1) ruvido (granulare), contenente ribosomi sulla sua superficie, e 2) liscio (agranulare), le cui membrane non portano ribosomi.
Funzioni: 1) trasporto di sostanze da una parte all'altra della cellula, 2) divisione del citoplasma cellulare in compartimenti ("compartimenti"), 3) sintesi di carboidrati e lipidi (RE liscio), 4) sintesi proteica (RE ruvido), 5) luogo di formazione dell'apparato di Golgi.
O Complesso di Golgi, è un organello a membrana singola. È costituito da pile di “cisterne” appiattite con bordi allargati. Ad essi è associato un sistema di piccole vescicole a membrana singola (vescicole di Golgi). Ogni pila è composta solitamente da 4-6 “cisterne”, è un'unità strutturale e funzionale dell'apparato di Golgi ed è chiamata dictiosoma. Il numero di dictosomi in una cellula varia da uno a diverse centinaia. Nelle cellule vegetali i dictosomi sono isolati.
L'apparato di Golgi si trova solitamente vicino al nucleo cellulare (nelle cellule animali, spesso vicino al centro cellulare).
Funzioni dell'apparato del Golgi: 1) accumulo di proteine, lipidi, carboidrati, 2) modifica delle sostanze organiche in entrata, 3) “impacchettamento” di proteine, lipidi, carboidrati in vescicole di membrana, 4) secrezione di proteine, lipidi, carboidrati, 5) sintesi di carboidrati e lipidi , 6) luogo di formazione dei lisosomi La funzione secretoria è la più importante, quindi l'apparato del Golgi è ben sviluppato nelle cellule secretorie.
Lisosomi
Lisosomi- organelli a membrana singola. Sono piccole bolle (diametro da 0,2 a 0,8 micron) contenenti un insieme di enzimi idrolitici. Gli enzimi vengono sintetizzati nel RE grezzo e si spostano nell'apparato di Golgi, dove vengono modificati e impacchettati in vescicole di membrana che, dopo la separazione dall'apparato di Golgi, diventano essi stessi lisosomi. Un lisosoma può contenere da 20 a 60 tipi diversi di enzimi idrolitici. Viene chiamata la scomposizione delle sostanze mediante enzimi lisi.
Ci sono: 1) lisosomi primari, 2) lisosomi secondari. Primari sono chiamati lisosomi che si staccano dall'apparato di Golgi. I lisosomi primari sono un fattore che garantisce l'esocitosi degli enzimi dalla cellula.
I secondari sono chiamati lisosomi formati come risultato della fusione dei lisosomi primari con vacuoli endocitici. In questo caso digeriscono le sostanze che entrano nella cellula attraverso la fagocitosi o la pinocitosi, per questo possono essere chiamati vacuoli digestivi.
Autofagia- il processo di distruzione delle strutture non necessarie per la cellula. Innanzitutto, la struttura da distruggere viene circondata da un'unica membrana, quindi la capsula di membrana risultante si fonde con il lisosoma primario, dando luogo alla formazione di un lisosoma secondario (vacuolo autofagico), in cui questa struttura viene digerita. I prodotti della digestione vengono assorbiti dal citoplasma cellulare, ma parte del materiale rimane non digerito. Il lisosoma secondario contenente questo materiale non digerito è chiamato corpo residuo. Per esocitosi, le particelle non digerite vengono rimosse dalla cellula.
Autolisi- autodistruzione cellulare, che avviene a causa del rilascio del contenuto dei lisosomi. Normalmente, l'autolisi avviene durante la metamorfosi (scomparsa della coda in un girino di rane), l'involuzione dell'utero dopo il parto e nelle aree di necrosi dei tessuti.
Funzioni dei lisosomi: 1) digestione intracellulare di sostanze organiche, 2) distruzione di strutture cellulari e non cellulari non necessarie, 3) partecipazione ai processi di riorganizzazione cellulare.
Vacuoli
Vacuoli- Gli organelli a membrana singola sono “contenitori” riempiti con soluzioni acquose di sostanze organiche e inorganiche. Il RE e l'apparato del Golgi partecipano alla formazione dei vacuoli. Le cellule giovani delle piante contengono molti piccoli vacuoli che poi, man mano che le cellule crescono e si differenziano, si fondono tra loro e formano un unico grande vacuolo centrale. Il vacuolo centrale può occupare fino al 95% del volume di una cellula matura; il nucleo e gli organelli vengono spinti verso la membrana cellulare. La membrana che delimita il vacuolo della pianta è chiamata tonoplasto. Si chiama il fluido che riempie il vacuolo di una pianta citoplasma. La composizione della linfa cellulare comprende sali organici e inorganici idrosolubili, monosaccaridi, disaccaridi, aminoacidi, prodotti metabolici finali o tossici (glicosidi, alcaloidi) e alcuni pigmenti (antociani).
Le cellule animali contengono piccoli vacuoli digestivi e autofagici, che appartengono al gruppo dei lisosomi secondari e contengono enzimi idrolitici. Gli animali unicellulari possiedono anche vacuoli contrattili che svolgono la funzione di osmoregolazione ed escrezione.
Funzioni del vacuolo: 1) accumulo e stoccaggio dell'acqua, 2) regolazione del metabolismo del sale marino, 3) mantenimento della pressione del turgore, 4) accumulo di metaboliti idrosolubili, sostanze nutritive di riserva, 5) colorazione di fiori e frutti e quindi attrazione di impollinatori e dispersori di semi , 6) vedi funzioni dei lisosomi.
Si formano il reticolo endoplasmatico, l'apparato di Golgi, i lisosomi e i vacuoli unica rete vacuolare della cellula, i cui singoli elementi possono trasformarsi l'uno nell'altro.
Mitocondri

1 - membrana esterna;
2 - membrana interna; 3 - matrice; 4 - cresta; 5 - sistema multienzimatico; 6 - DNA circolare.
La forma, la dimensione e il numero dei mitocondri variano enormemente. I mitocondri possono essere a forma di bastoncino, rotondi, a spirale, a forma di coppa o ramificati. La lunghezza dei mitocondri varia da 1,5 a 10 µm, il diametro da 0,25 a 1,00 µm. Il numero di mitocondri in una cellula può raggiungere diverse migliaia e dipende dall'attività metabolica della cellula.
Il mitocondrio è delimitato da due membrane. La membrana esterna dei mitocondri (1) è liscia, quella interna (2) forma numerose pieghe - cristas(4). Le cristae aumentano la superficie della membrana interna, sulla quale sono localizzati sistemi multienzimatici (5) coinvolti nella sintesi delle molecole di ATP. Lo spazio interno dei mitocondri è pieno di matrice (3). La matrice contiene DNA circolare (6), mRNA specifico, ribosomi di tipo procariotico (tipo 70S) ed enzimi del ciclo di Krebs.
Il DNA mitocondriale non è associato alle proteine (“nudo”), è attaccato alla membrana interna del mitocondrio e trasporta informazioni sulla struttura di circa 30 proteine. Per costruire un mitocondrio sono necessarie molte più proteine, quindi le informazioni sulla maggior parte delle proteine mitocondriali sono contenute nel DNA nucleare e queste proteine sono sintetizzate nel citoplasma della cellula. I mitocondri sono capaci di riprodursi autonomamente mediante fissione in due. Tra la membrana esterna e quella interna c'è serbatoio di protoni, dove avviene l'accumulo di H+.
Funzioni dei mitocondri: 1) Sintesi di ATP, 2) degradazione dell'ossigeno delle sostanze organiche.
Secondo un'ipotesi (la teoria della simbiogenesi), i mitocondri hanno avuto origine da antichi organismi procarioti aerobici a vita libera che, essendo penetrati accidentalmente nella cellula ospite, hanno poi formato con essa un complesso simbiotico reciprocamente vantaggioso. I seguenti dati supportano questa ipotesi. Innanzitutto, il DNA mitocondriale ha le stesse caratteristiche strutturali del DNA dei batteri moderni (chiuso ad anello, non associato a proteine). In secondo luogo, i ribosomi mitocondriali e i ribosomi batterici appartengono allo stesso tipo: il tipo 70S. In terzo luogo, il meccanismo della fissione mitocondriale è simile a quello dei batteri. In quarto luogo, la sintesi delle proteine mitocondriali e batteriche viene soppressa dagli stessi antibiotici.
Plastidi

1 - membrana esterna; 2 - membrana interna; 3 - stroma; 4 - tilacoide; 5 - grana; 6 - lamelle; 7 - grani di amido; 8 - gocce lipidiche.
I plastidi sono caratteristici solo delle cellule vegetali. Distinguere tre tipi principali di plastidi: i leucoplasti sono plastidi incolori presenti nelle cellule di parti incolori delle piante, i cromoplasti sono plastidi colorati solitamente gialli, rossi e arancioni, i cloroplasti sono plastidi verdi.
Cloroplasti. Nelle cellule delle piante superiori, i cloroplasti hanno la forma di una lente biconvessa. La lunghezza dei cloroplasti varia da 5 a 10 µm, il diametro da 2 a 4 µm. I cloroplasti sono delimitati da due membrane. La membrana esterna (1) è liscia, quella interna (2) ha una struttura piegata complessa. Si chiama la piega più piccola tilacoide(4). Viene chiamato un gruppo di tilacoidi disposti come una pila di monete sfaccettatura(5). Il cloroplasto contiene mediamente 40-60 grani, disposti a scacchiera. Le granae sono collegate tra loro da canali appiattiti - lamelle(6). Le membrane tilacoidi contengono pigmenti fotosintetici ed enzimi che forniscono la sintesi di ATP. Il principale pigmento fotosintetico è la clorofilla, che determina il colore verde dei cloroplasti.
Lo spazio interno dei cloroplasti è pieno stroma(3). Lo stroma contiene DNA circolare “nudo”, ribosomi di tipo 70S, enzimi del ciclo di Calvin e granuli di amido (7). All'interno di ciascun tilacoide c'è un serbatoio di protoni e si accumula H +. I cloroplasti, come i mitocondri, sono capaci di riprodursi autonomamente dividendosi in due. Si trovano nelle cellule delle parti verdi delle piante superiori, in particolare in molti cloroplasti nelle foglie e nei frutti verdi. I cloroplasti delle piante inferiori sono detti cromatofori.
Funzione dei cloroplasti: fotosintesi. Si ritiene che i cloroplasti provengano da antichi cianobatteri endosimbiotici (teoria della simbiogenesi). La base di questa ipotesi è la somiglianza dei cloroplasti e dei batteri moderni in una serie di caratteristiche (DNA circolare, “nudo”, ribosomi di tipo 70S, metodo di riproduzione).
Leucoplasti. La forma varia (sferica, rotonda, a coppa, ecc.). I leucoplasti sono delimitati da due membrane. La membrana esterna è liscia, quella interna forma pochi tilacoidi. Lo stroma contiene DNA circolare “nudo”, ribosomi di tipo 70S, enzimi per la sintesi e l'idrolisi dei nutrienti di riserva. Non ci sono pigmenti. Le cellule degli organi sotterranei della pianta (radici, tuberi, rizomi, ecc.) possiedono soprattutto numerosi leucoplasti. Funzione dei leucoplasti: sintesi, accumulo e stoccaggio dei nutrienti di riserva. Amiloplasti- leucoplasti che sintetizzano e accumulano amido, elaioplasti- oli, proteinaplasti- proteine. Nello stesso leucoplasto possono accumularsi sostanze diverse.
Cromoplasti. Delimitato da due membrane. La membrana esterna è liscia, la membrana interna è liscia o forma singoli tilacoidi. Lo stroma contiene DNA circolare e pigmenti: carotenoidi, che conferiscono ai cromoplasti un colore giallo, rosso o arancione. La forma di accumulo dei pigmenti è diversa: sotto forma di cristalli, disciolti in goccioline lipidiche (8), ecc. Contenuto nelle cellule di frutti maturi, petali, foglie autunnali e raramente - ortaggi a radice. I cromoplasti sono considerati lo stadio finale dello sviluppo dei plastidi.
Funzione dei cromoplasti: colorando fiori e frutti e attirando così gli impollinatori e i dispersori di semi.
Tutti i tipi di plastidi possono essere formati da proplastidi. Proplastidi- piccoli organelli contenuti nei tessuti meristematici. Poiché i plastidi hanno un'origine comune, sono possibili interconversioni tra loro. I leucoplasti possono trasformarsi in cloroplasti (inverdimento dei tuberi di patata alla luce), cloroplasti - in cromoplasti (ingiallimento delle foglie e arrossamento dei frutti). La trasformazione dei cromoplasti in leucoplasti o cloroplasti è considerata impossibile.
Ribosomi

1 - subunità grande; 2 - piccola subunità.
Ribosomi- organelli non di membrana, diametro di circa 20 nm. I ribosomi sono costituiti da due subunità: grande e piccola, nelle quali possono dissociarsi. La composizione chimica dei ribosomi è costituita da proteine e rRNA. Le molecole di rRNA costituiscono il 50-63% della massa del ribosoma e ne formano la struttura strutturale. Esistono due tipi di ribosomi: 1) eucariotici (con costanti di sedimentazione per l'intero ribosoma - 80S, subunità piccola - 40S, grande - 60S) e 2) procariotici (70S, 30S, 50S, rispettivamente).
I ribosomi di tipo eucariotico contengono 4 molecole di rRNA e circa 100 molecole di proteine, mentre il tipo procariotico contiene 3 molecole di rRNA e circa 55 molecole di proteine. Durante la biosintesi delle proteine, i ribosomi possono “lavorare” individualmente o combinarsi in complessi - poliribosomi (polisomi). In tali complessi sono collegati tra loro da una molecola di mRNA. Le cellule procariotiche hanno solo ribosomi di tipo 70S. Le cellule eucariotiche hanno ribosomi sia di tipo 80S (membrane EPS ruvide, citoplasma) che di tipo 70 (mitocondri, cloroplasti).
Le subunità ribosomiali eucariotiche si formano nel nucleolo. La combinazione delle subunità in un intero ribosoma avviene nel citoplasma, solitamente durante la biosintesi delle proteine.
Funzione dei ribosomi: assemblaggio di una catena polipeptidica (sintesi proteica).
Citoscheletro
Citoscheletro formato da microtubuli e microfilamenti. I microtubuli sono strutture cilindriche e non ramificate. La lunghezza dei microtubuli varia da 100 µm a 1 mm, il diametro è di circa 24 nm e lo spessore della parete è di 5 nm. Il principale componente chimico è la tubulina proteica. I microtubuli vengono distrutti dalla colchicina. I microfilamenti sono filamenti con un diametro di 5-7 nm e sono costituiti dalla proteina actina. Microtubuli e microfilamenti formano trame complesse nel citoplasma. Funzioni del citoscheletro: 1) determinazione della forma della cellula, 2) supporto per organelli, 3) formazione del fuso, 4) partecipazione ai movimenti cellulari, 5) organizzazione del flusso citoplasmatico.

Include due centrioli e una centrosfera. Centrioloè un cilindro, la cui parete è formata da nove gruppi di tre microtubuli fusi (9 triplette), interconnessi a determinati intervalli da legami incrociati. I centrioli sono uniti a coppie dove si trovano ad angolo retto tra loro. Prima della divisione cellulare, i centrioli divergono ai poli opposti e vicino a ciascuno di essi appare un centriolo figlio. Formano un fuso di divisione, che contribuisce alla distribuzione uniforme del materiale genetico tra le cellule figlie. Nelle cellule delle piante superiori (gimnosperme, angiosperme), il centro cellulare non ha centrioli. I centrioli sono organelli autoreplicanti del citoplasma; nascono come risultato della duplicazione dei centrioli esistenti. Funzioni: 1) garantendo la divergenza dei cromosomi rispetto ai poli cellulari durante la mitosi o la meiosi, 2) il centro di organizzazione del citoscheletro.
Organoidi del movimento
Non presente in tutte le cellule. Gli organelli di movimento includono ciglia (ciliati, epitelio delle vie respiratorie), flagelli (flagellati, sperma), pseudopodi (rizopodi, leucociti), miofibrille (cellule muscolari), ecc.
Flagelli e ciglia- organelli a forma di filamento, che rappresentano un assonema delimitato da una membrana. L'assonema è una struttura cilindrica; la parete del cilindro è formata da nove paia di microtubuli; al suo centro si trovano due microtubuli singoli. Alla base dell'assonema ci sono i corpi basali, rappresentati da due centrioli tra loro perpendicolari (ciascun corpo basale è costituito da nove triplette di microtubuli; al suo centro non ci sono microtubuli). La lunghezza del flagello raggiunge i 150 micron, le ciglia sono molte volte più corte.
Miofibrille sono costituiti da miofilamenti di actina e miosina che forniscono la contrazione delle cellule muscolari.
Vai a lezioni n. 6“Cellula eucariotica: citoplasma, membrana cellulare, struttura e funzioni delle membrane cellulari”